

DNA Polymerase Delta Synthesizes Both Strands during Break-Induced Replication
- PMID: 31495565
- PMCID: PMC6862718
- DOI: 10.1016/j.molcel.2019.07.033
DNA Polymerase Delta Synthesizes Both Strands during Break-Induced Replication
Abstract
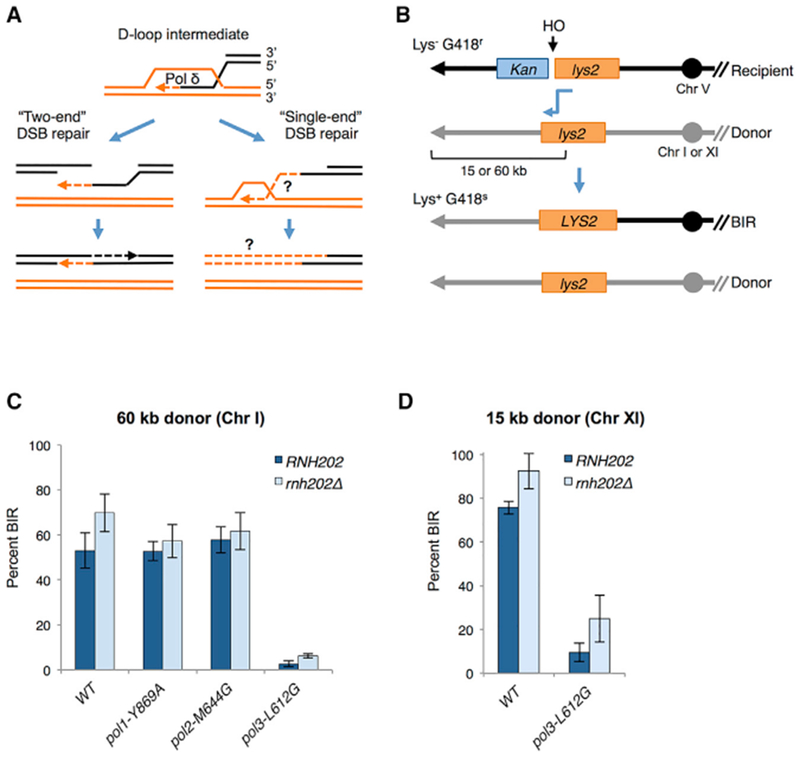
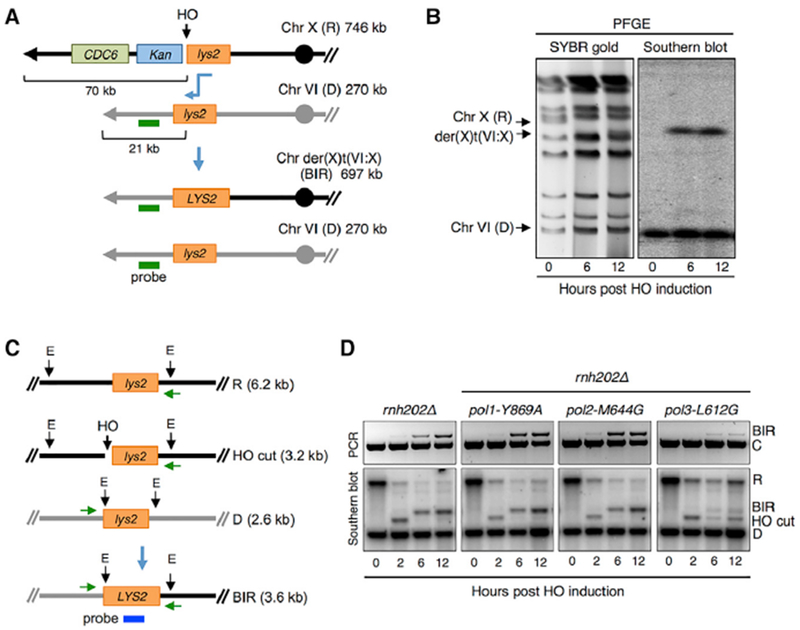
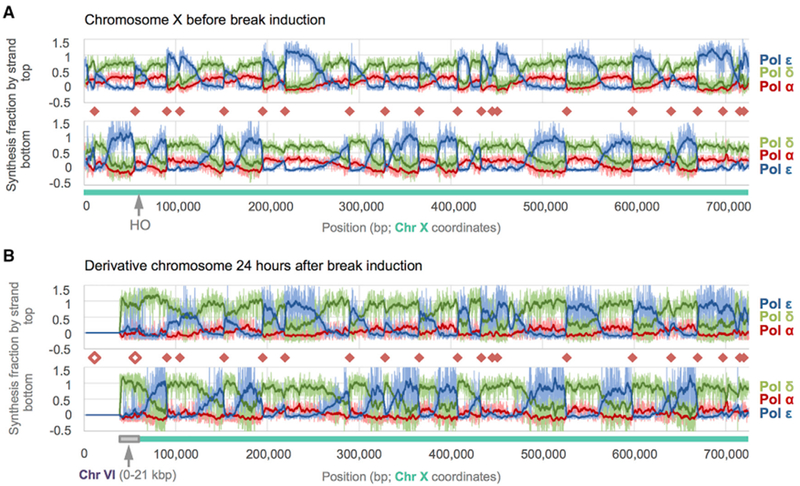
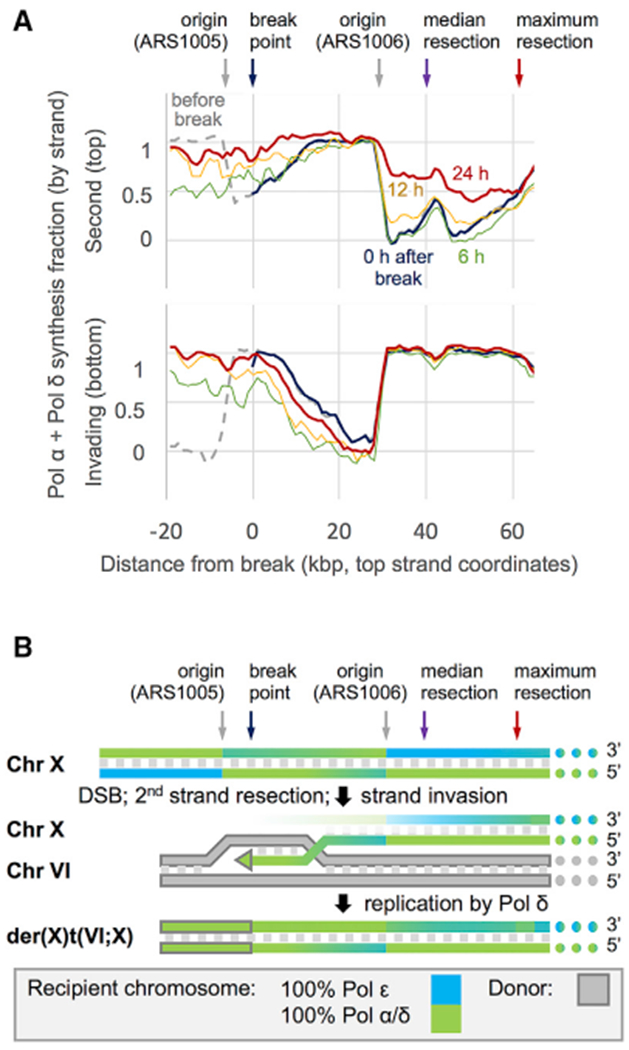
Break-induced replication (BIR) is a pathway of homology-directed repair that repairs one-ended DNA breaks, such as those formed at broken replication forks or uncapped telomeres. In contrast to conventional S phase DNA synthesis, BIR proceeds by a migrating D-loop and results in conservative synthesis of the nascent strands. DNA polymerase delta (Pol δ) initiates BIR; however, it is not known whether synthesis of the invading strand switches to a different polymerase or how the complementary strand is synthesized. By using alleles of the replicative DNA polymerases that are permissive for ribonucleotide incorporation, thus generating a signature of their action in the genome that can be identified by hydrolytic end sequencing, we show that Pol δ replicates both the invading and the complementary strand during BIR. In support of this conclusion, we show that depletion of Pol δ from cells reduces BIR, whereas depletion of Pol ε has no effect.
Keywords: DNA polymerase delta; DNA repair; break-induced replication; homologous recombination.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
SUPPLEMENTAL INFORMATION
Supplemental Information can be found online at
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
References
-
- Amberg DC, Burke DJ, and Strathern JN (2005). Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual (Cold Spring Harbor Laboratory Press; ).
-
- Bhowmick R, Minocherhomji S, and Hickson ID (2016). RAD52 facilitates mitotic DNA synthesis following replication stress. Mol. Cell 64, 1117–1126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
