

Gut-Proglucagon-Derived Peptides Are Essential for Regulating Glucose Homeostasis in Mice
- PMID: 31495689
- PMCID: PMC8140521
- DOI: 10.1016/j.cmet.2019.08.009
Gut-Proglucagon-Derived Peptides Are Essential for Regulating Glucose Homeostasis in Mice
Abstract
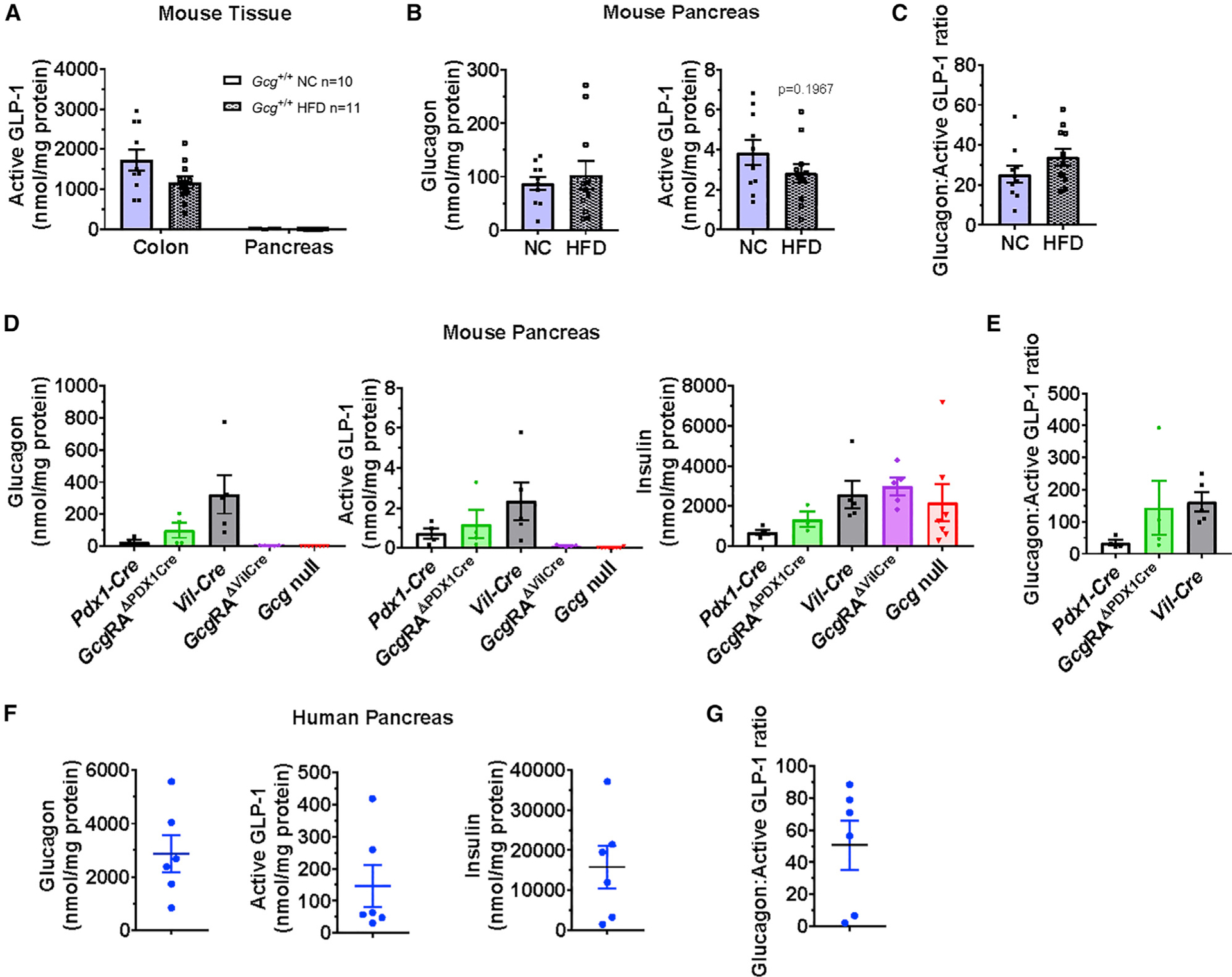
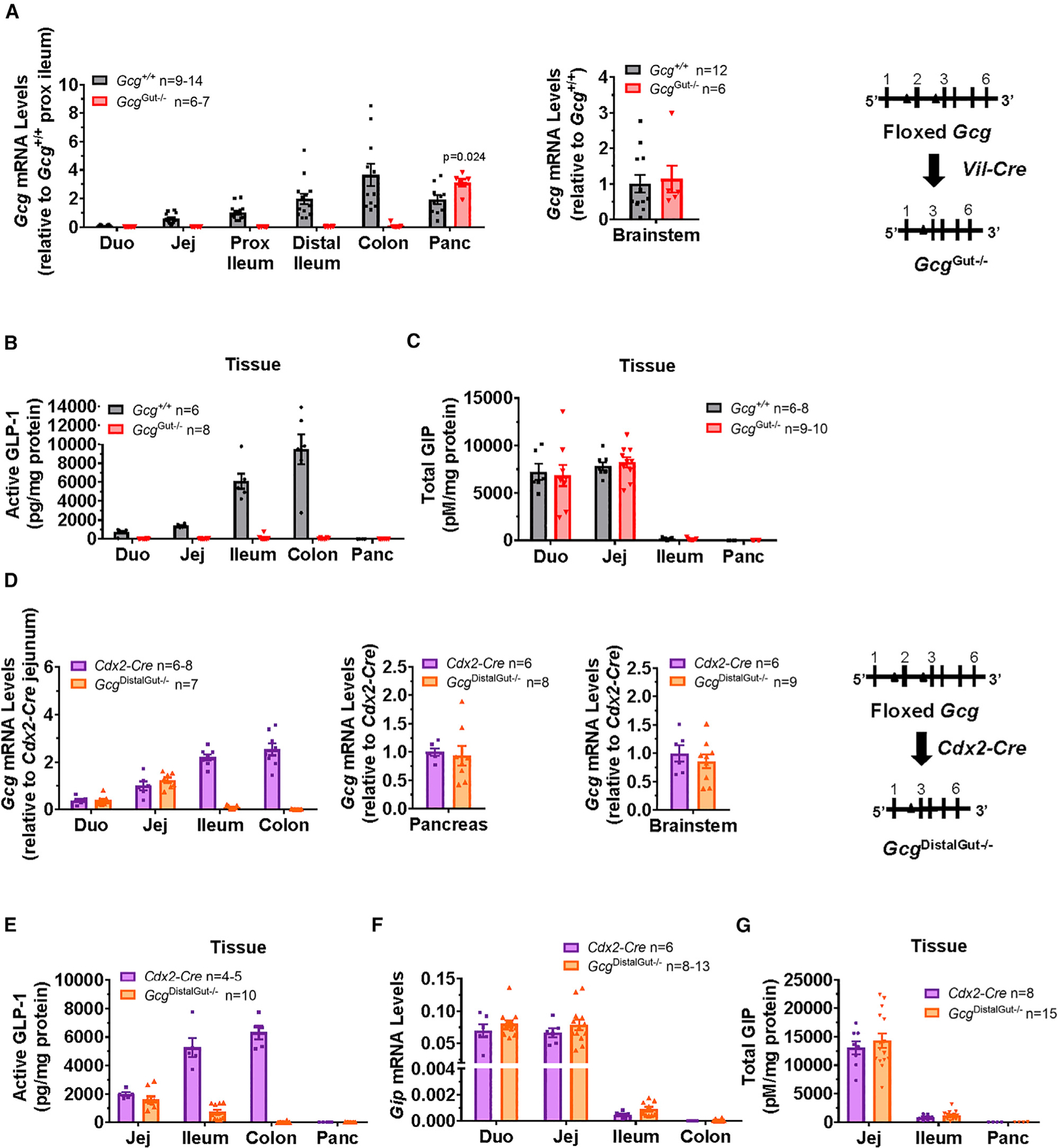
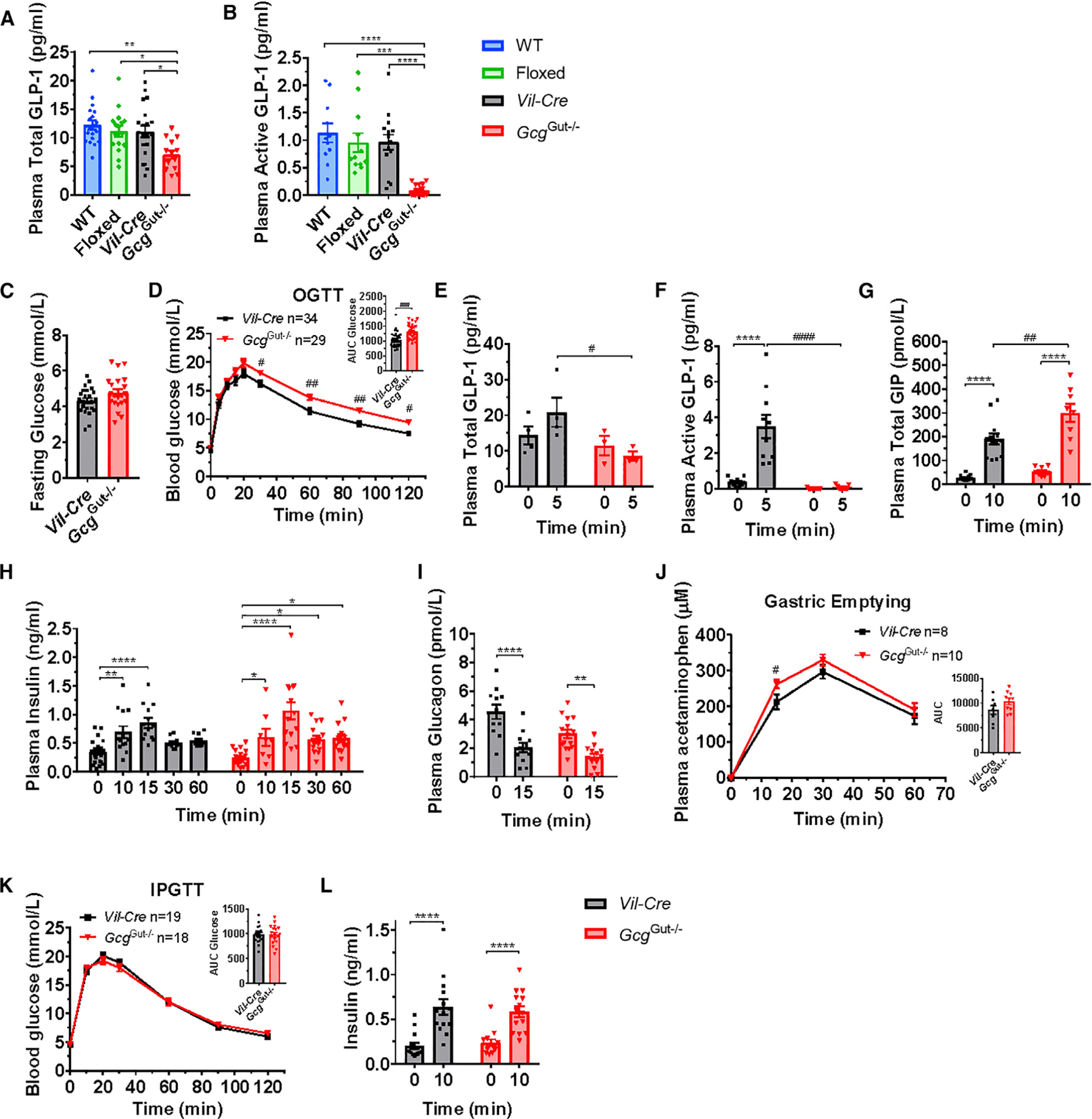
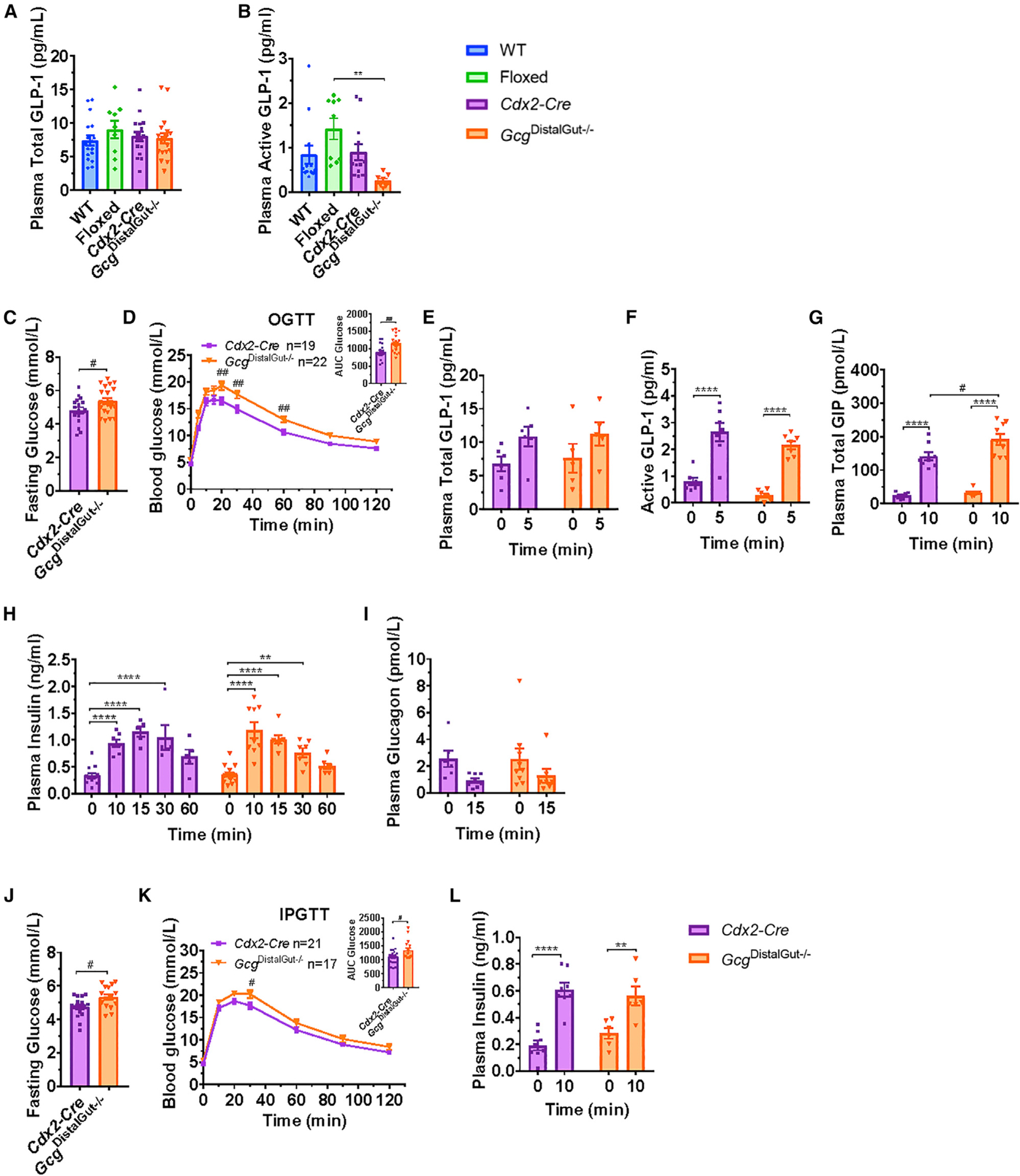
The importance of pancreatic versus intestinal-derived GLP-1 for glucose homeostasis is controversial. We detected active GLP-1 in the mouse and human pancreas, albeit at extremely low levels relative to glucagon. Accordingly, to elucidate the metabolic importance of intestinal proglucagon-derived peptides (PGDPs), we generated mice with reduction of Gcg expression within the distal (GcgDistalGut-/-) or entire (GcgGut-/-) gut. Substantial reduction of gut Gcg expression markedly reduced circulating levels of GLP-1, and impaired glucose homeostasis, associated with increased levels of GIP, and accelerated gastric emptying. GcgDistalGut-/- mice similarly exhibited lower circulating GLP-1 and impaired oral glucose tolerance. Nevertheless, plasma levels of insulin remained normal following glucose administration in the absence of gut-derived GLP-1. Collectively, our findings identify the essential importance of gut-derived PGDPs for maintaining levels of circulating GLP-1, control of gastric emptying, and glucose homeostasis.
Keywords: GIP; GLP-1; diabetes; enteroendocrine cell; glucagon; incretin; insulin; intestine; islet; pancreas.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
D.J.D. has served as an advisor, consultant, or speaker in the past 12 months to Forkhead Biotherapeutics, Heliome, Inc., Intarcia Therapeutics, Kallyope, Eli Lilly, Merck Research Laboratories, Novo Nordisk, Inc., Pfizer, Inc., and Sanofi, Inc. Neither D.J.D. nor his family members hold stock directly or indirectly in any of these companies.
Figures
Comment on
-
The Role of Pancreatic Preproglucagon in Glucose Homeostasis in Mice.Cell Metab. 2017 Apr 4;25(4):927-934.e3. doi: 10.1016/j.cmet.2017.02.008. Epub 2017 Mar 16. Cell Metab. 2017. PMID: 28325479 Free PMC article.
References
-
- Campbell JE, and Drucker DJ (2013). Pharmacology physiology and mechanisms of incretin hormone action. Cell Metab 17, 819–837. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
