

Cohesin Members Stag1 and Stag2 Display Distinct Roles in Chromatin Accessibility and Topological Control of HSC Self-Renewal and Differentiation
- PMID: 31495782
- PMCID: PMC6842438
- DOI: 10.1016/j.stem.2019.08.003
Cohesin Members Stag1 and Stag2 Display Distinct Roles in Chromatin Accessibility and Topological Control of HSC Self-Renewal and Differentiation
Abstract
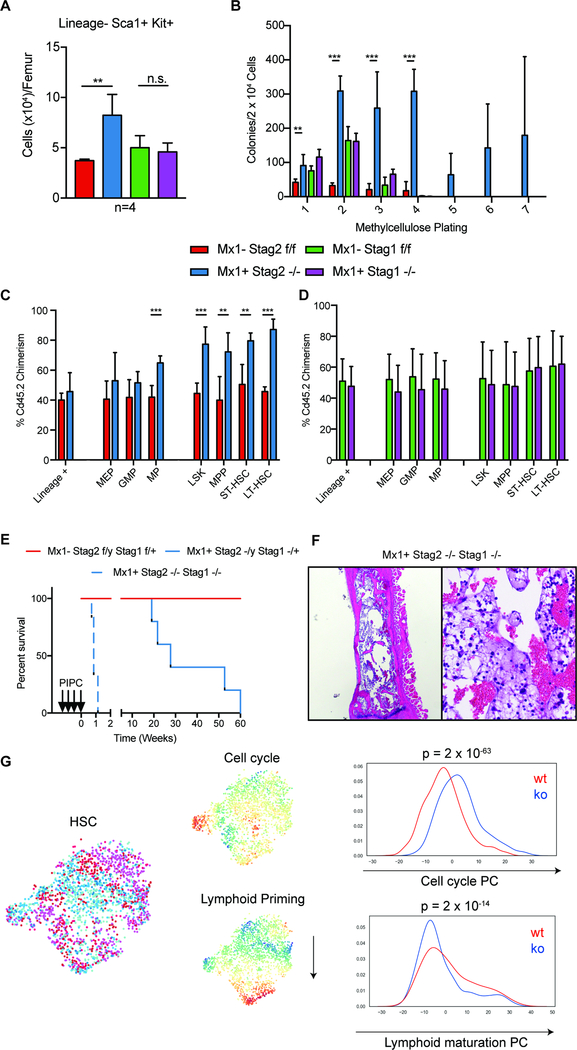
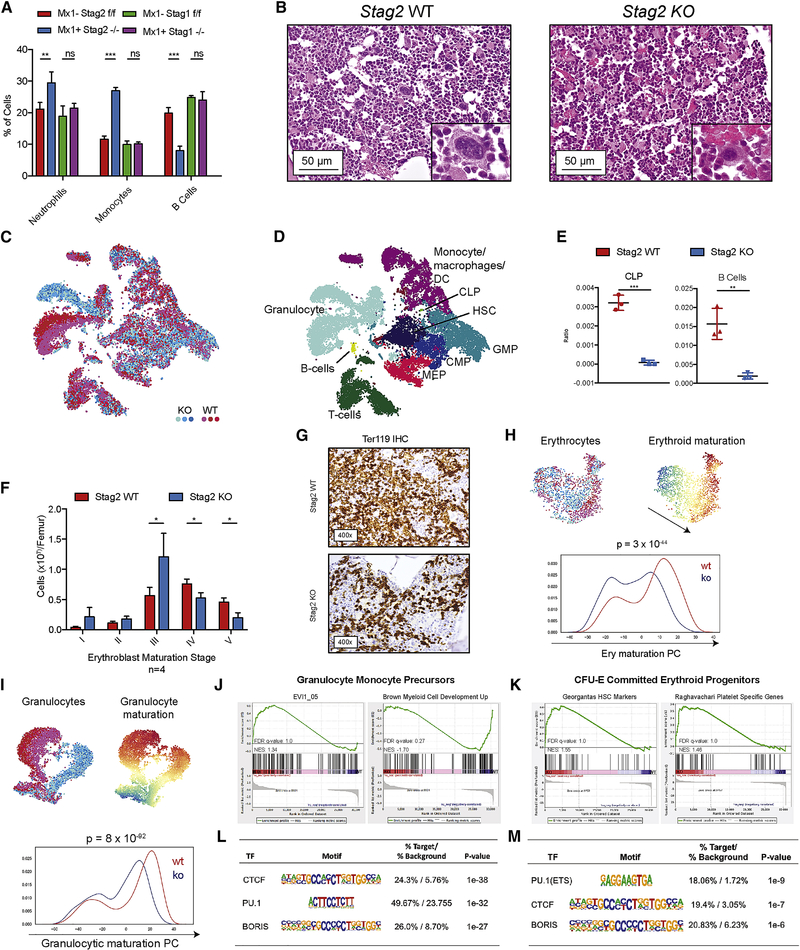
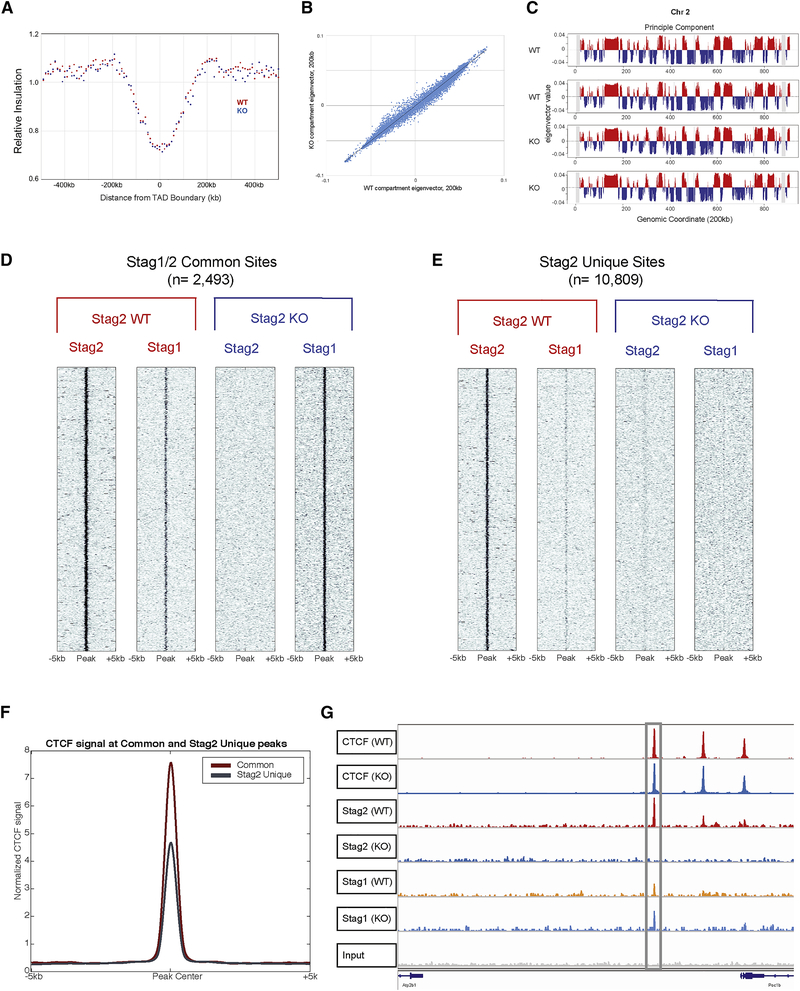
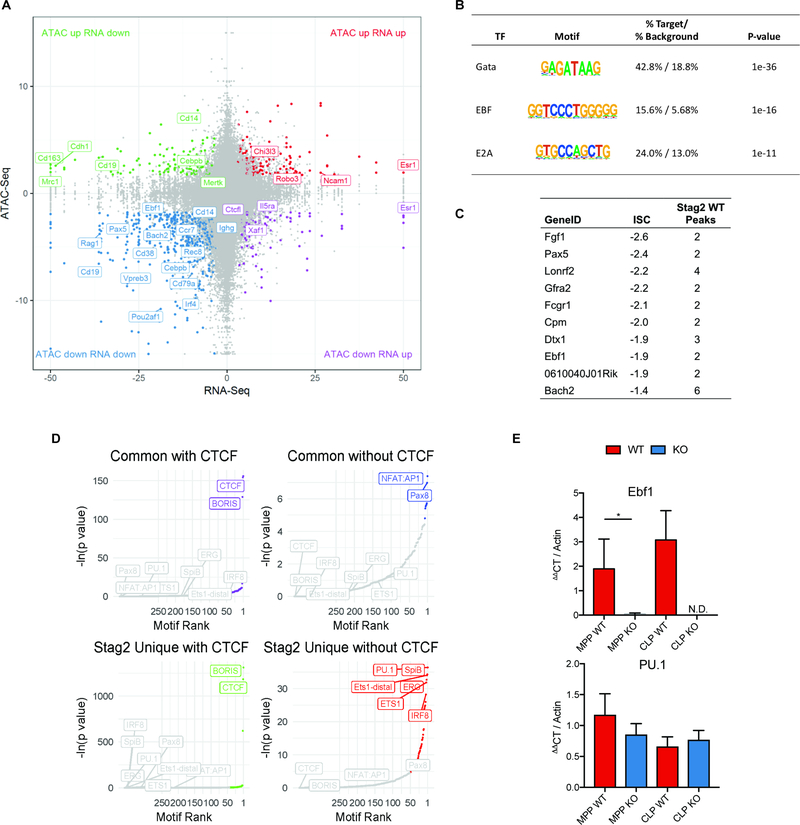
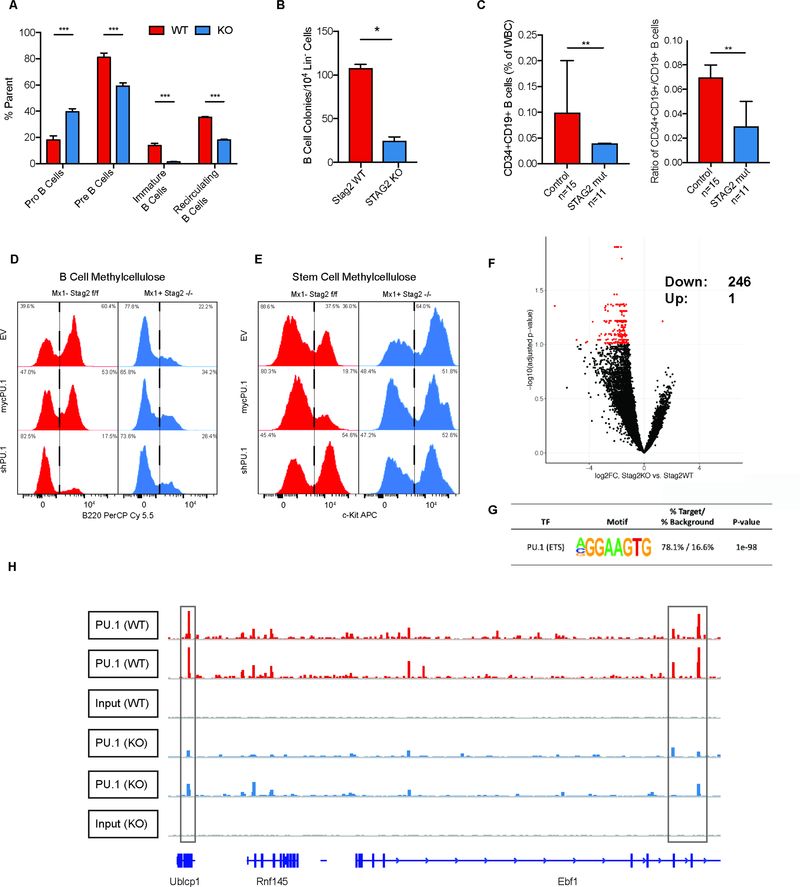
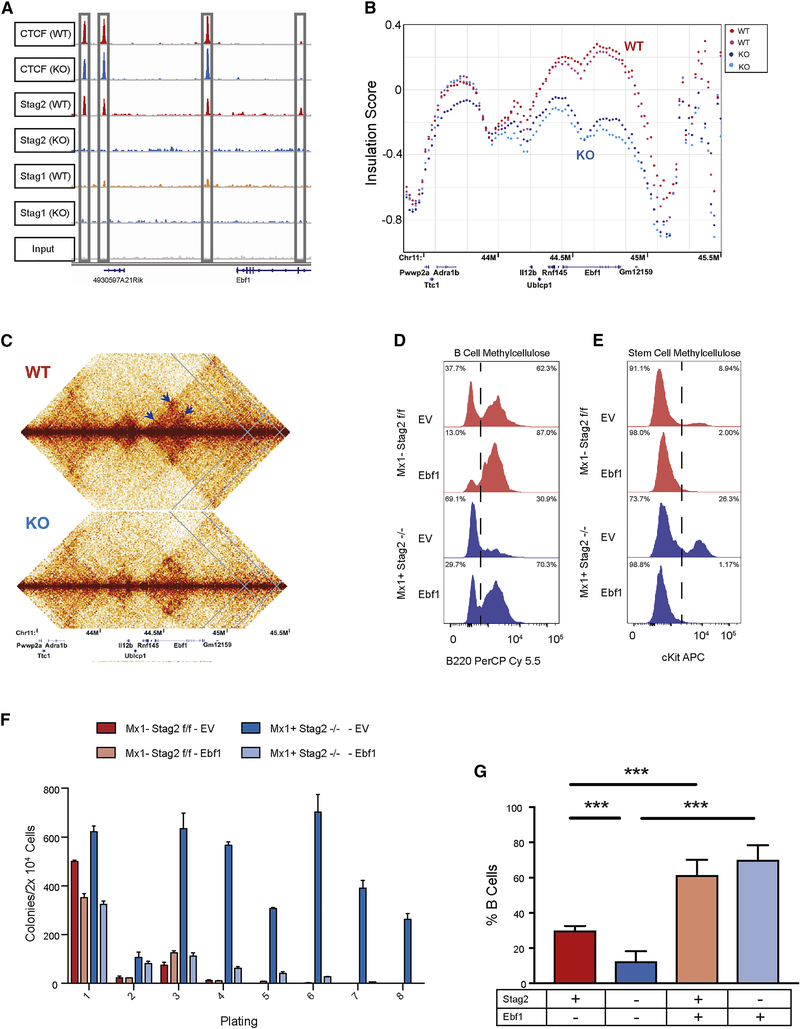
Transcriptional regulators, including the cohesin complex member STAG2, are recurrently mutated in cancer. The role of STAG2 in gene regulation, hematopoiesis, and tumor suppression remains unresolved. We show that Stag2 deletion in hematopoietic stem and progenitor cells (HSPCs) results in altered hematopoietic function, increased self-renewal, and impaired differentiation. Chromatin immunoprecipitation (ChIP) sequencing revealed that, although Stag2 and Stag1 bind a shared set of genomic loci, a component of Stag2 binding sites is unoccupied by Stag1, even in Stag2-deficient HSPCs. Although concurrent loss of Stag2 and Stag1 abrogated hematopoiesis, Stag2 loss alone decreased chromatin accessibility and transcription of lineage-specification genes, including Ebf1 and Pax5, leading to increased self-renewal and reduced HSPC commitment to the B cell lineage. Our data illustrate a role for Stag2 in transformation and transcriptional dysregulation distinct from its shared role with Stag1 in chromosomal segregation.
Keywords: Cohesin; Stag1; Stag2; chromatin; hematopoietic stem cells; mouse models; myelodysplasia; nuclear topology.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures
References
-
- Adolfsson J, Borge OJ, Bryder D, Theilgaard-Monch K, Astrand-Grundstrom I, Sitnicka E, Sasaki Y, and Jacobsen SE (2001). Upregulation of Flt3 expression within the bone marrow Lin(−)Sca1(+)c-kit(+) stem cell compartment is accompanied by loss of self-renewal capacity. Immunity 15, 659–669. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
