

Whisker-Mediated Touch System in Rodents: From Neuron to Behavior
- PMID: 31496942
- PMCID: PMC6712080
- DOI: 10.3389/fnsys.2019.00040
Whisker-Mediated Touch System in Rodents: From Neuron to Behavior
Abstract
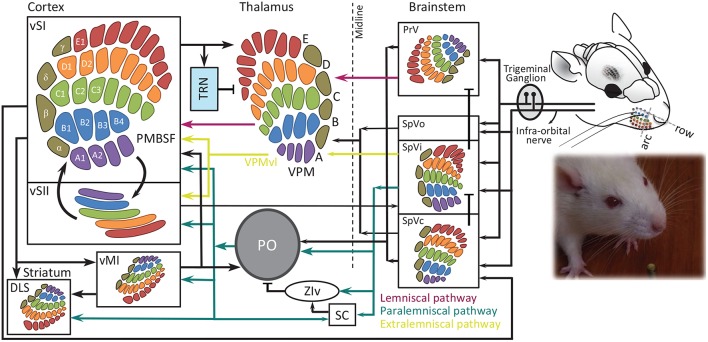
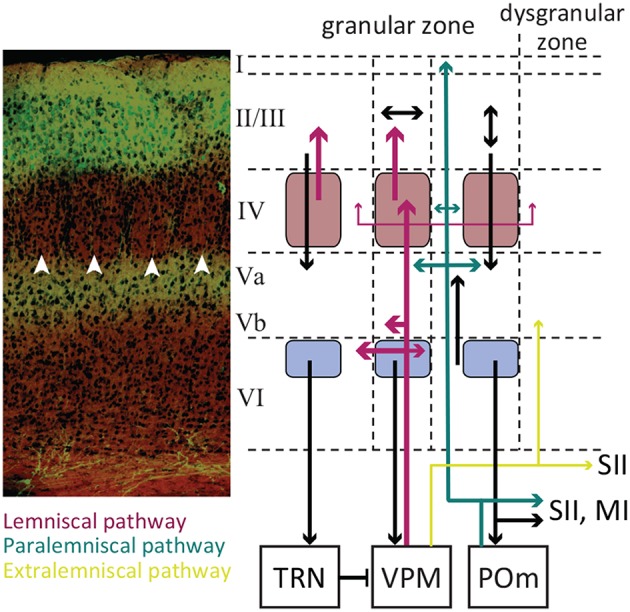
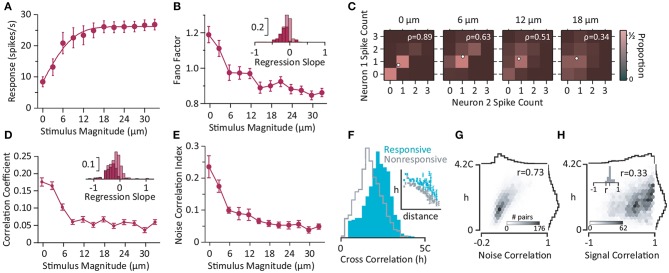
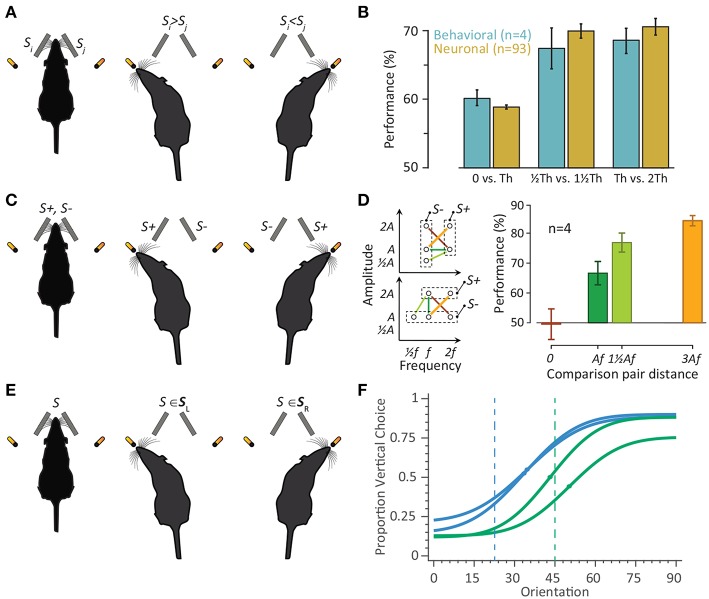
A key question in systems neuroscience is to identify how sensory stimuli are represented in neuronal activity, and how the activity of sensory neurons in turn is "read out" by downstream neurons and give rise to behavior. The choice of a proper model system to address these questions, is therefore a crucial step. Over the past decade, the increasingly powerful array of experimental approaches that has become available in non-primate models (e.g., optogenetics and two-photon imaging) has spurred a renewed interest for the use of rodent models in systems neuroscience research. Here, I introduce the rodent whisker-mediated touch system as a structurally well-established and well-organized model system which, despite its simplicity, gives rise to complex behaviors. This system serves as a behaviorally efficient model system; known as nocturnal animals, along with their olfaction, rodents rely on their whisker-mediated touch system to collect information about their surrounding environment. Moreover, this system represents a well-studied circuitry with a somatotopic organization. At every stage of processing, one can identify anatomical and functional topographic maps of whiskers; "barrelettes" in the brainstem nuclei, "barreloids" in the sensory thalamus, and "barrels" in the cortex. This article provides a brief review on the basic anatomy and function of the whisker system in rodents.
Keywords: barrel field; rodents; somatosensory; thalamic barreloids; vibrissae; vibrissal system; whisker system.
Figures
References
-
- Adelson E. H., Bergen J. R. (1985). Spatiotemporal energy models for the perception of motion. J. Opt. Soc. Am. A 2, 284–299. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous