

Cloning and identification of antimicrobial peptide, hepcidin from freshwater carp, Catla catla on pathogen challenge and PAMPs stimulation
- PMID: 31497459
- PMCID: PMC6707987
- DOI: 10.1007/s13205-019-1874-6
Cloning and identification of antimicrobial peptide, hepcidin from freshwater carp, Catla catla on pathogen challenge and PAMPs stimulation
Abstract
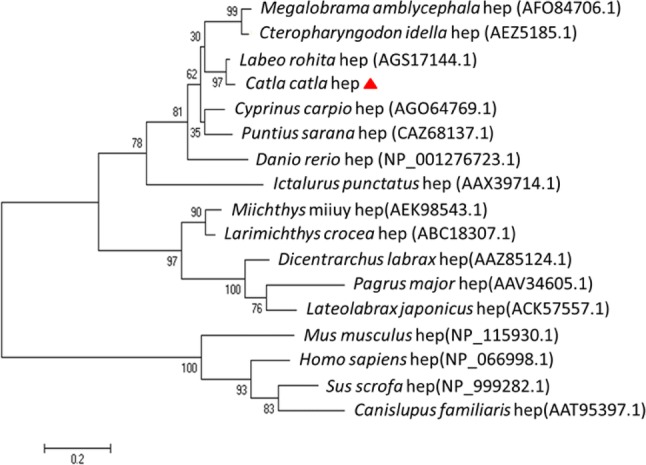
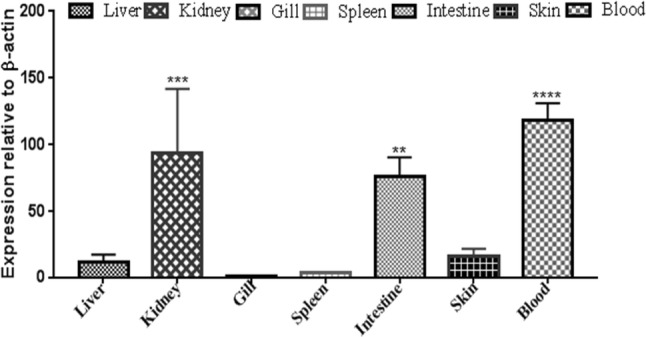
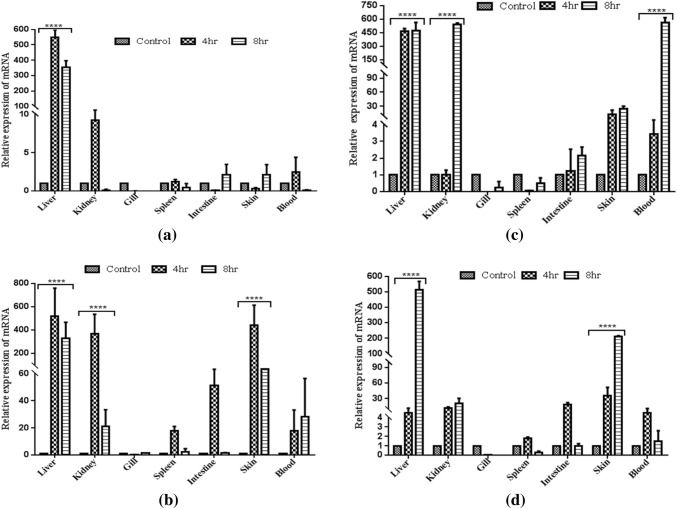
Hepcidin, a cationic cysteine-rich antimicrobial peptide (AMP) acts in hormone regulation and iron homeostasis in the host body. However, the biological property of hepcidin in immune reaction remains unexplored. In aquatic milieu, environmental and pathogenic stressors cause detrimental infections, which are defended by various immunological cells and antimicrobial peptides. In this study, hepcidin gene has been cloned from freshwater carp, Catla catla. The partially cloned hepcidin consists of 200 bp nucleotide sequence encoding 66 amino acids. Nucleotide sequence showed 97% and 91% similarity with Labeo rohita and Cyprinus carpio, respectively. Expression profile revealed significant up-regulation (P ≤ 0.0001) in liver as compared to other tissues in different conditions. In Aeromonas hydrophila challenged C. catla, liver showed higher expression level of hepcidin at 72 h as compared to other tissues. In skin, hepcidin expression showed significant upraise during 24 h in Streptococcus uberis infection. In Argulus sp. infected fishes, up-regulation of hepcidin expression was noted in liver, intestine and skin. The inactivated viral antigen-stimulated fishes, a substantial rise in liver was observed implying hepcidin as an important molecule in combating the pathogenic infections in freshwater carp, C. catla. Fishes stimulated with pathogen-associated molecular patterns (PAMPs) triggered the increased expression of hepcidin mRNA in liver, kidney and skin. This study indicates the presence of hepcidin as antimicrobial peptide in neutralizing the pathogenic infection in fishes.
Keywords: Antimicrobial peptide; Catla catla; Hepcidin; PAMPs; Pathogen challenge; Transcriptomic expression.
Conflict of interest statement
Conflict of interestThe authors declare no conflict of interest.
Figures





References
-
- Banerjee R, Patel B, Basu M, Lenka SS, Paicha M, Samanta M, Das S. Molecular cloning, characterization and expression of immunoglobulin D on pathogen challenge and pathogen associated molecular patterns stimulation in freshwater carp, Catla catla. Microbiol Immunol. 2017;61(10):452–458. doi: 10.1111/1348-0421.12534. - DOI - PubMed
-
- Basu M, Swain B, Maiti NK, Routray P, Samanta M. Inductive expression of toll-like receptor 5 (TLR5) and associated downstream signaling molecules following ligand exposure and bacterial infection in the Indian major carp, mrigal (Cirrhinus mrigala) Fish Shellfish Immunol. 2012;32(1):121–131. doi: 10.1016/j.fsi.2011.10.031. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
