

Wood Transcriptome Profiling Identifies Critical Pathway Genes of Secondary Wall Biosynthesis and Novel Regulators for Vascular Cambium Development in Populus
- PMID: 31500311
- PMCID: PMC6770981
- DOI: 10.3390/genes10090690
Wood Transcriptome Profiling Identifies Critical Pathway Genes of Secondary Wall Biosynthesis and Novel Regulators for Vascular Cambium Development in Populus
Abstract
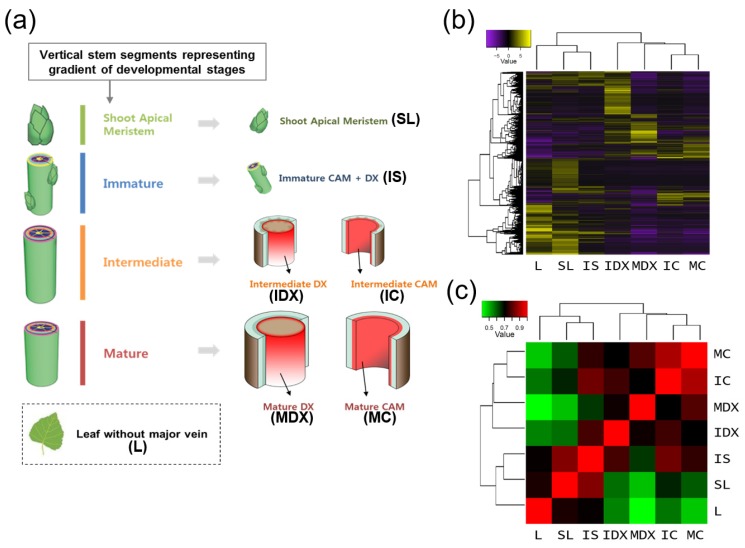
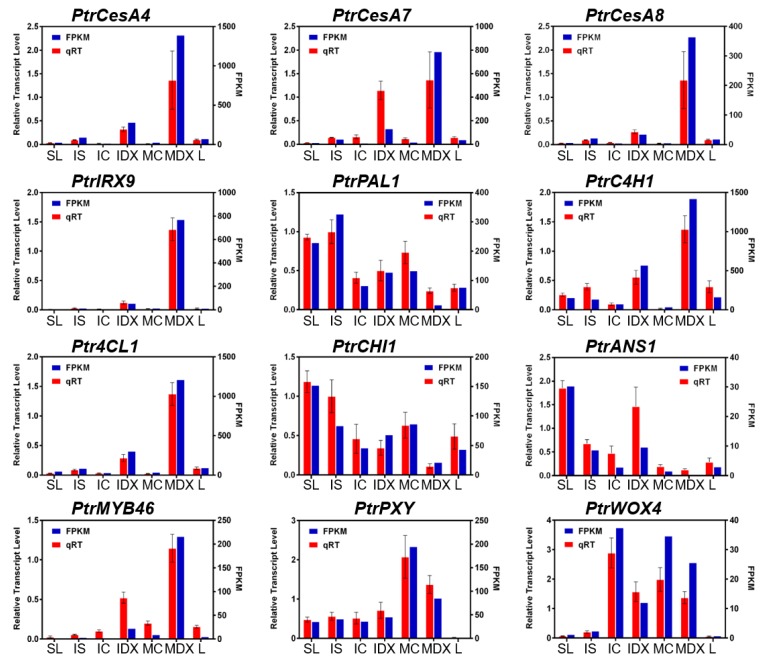
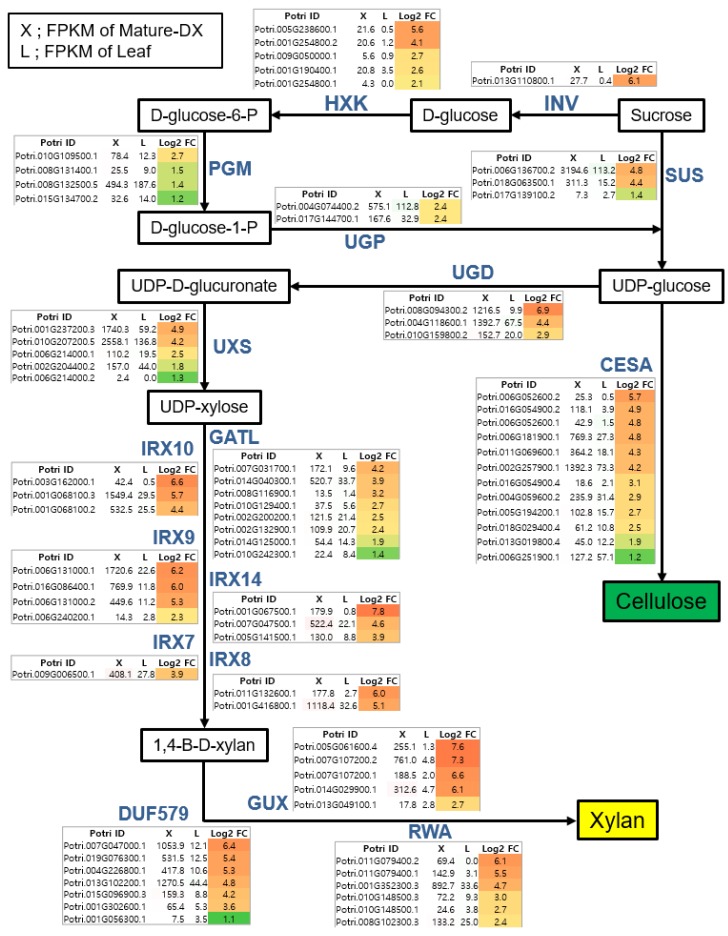
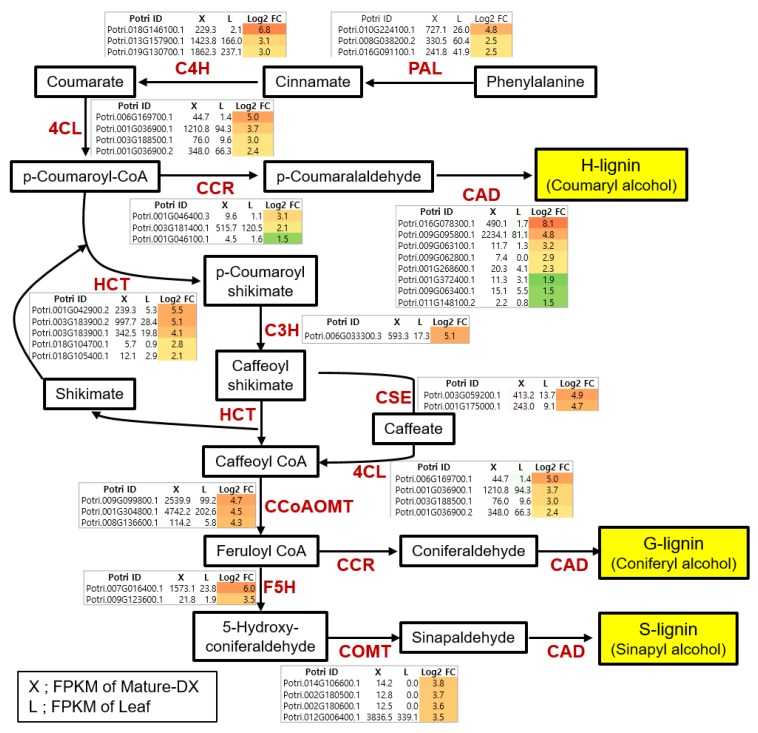
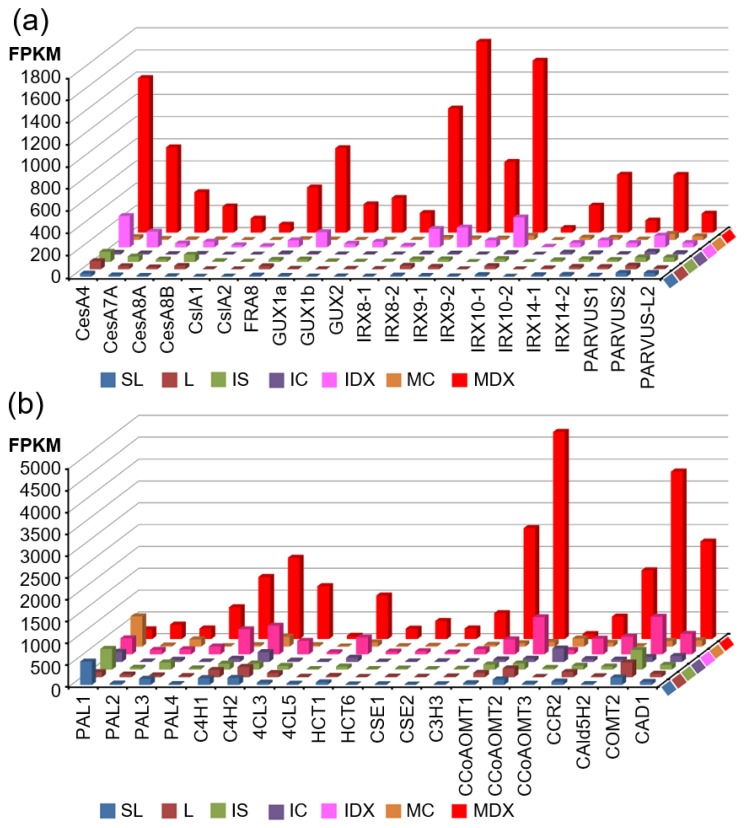
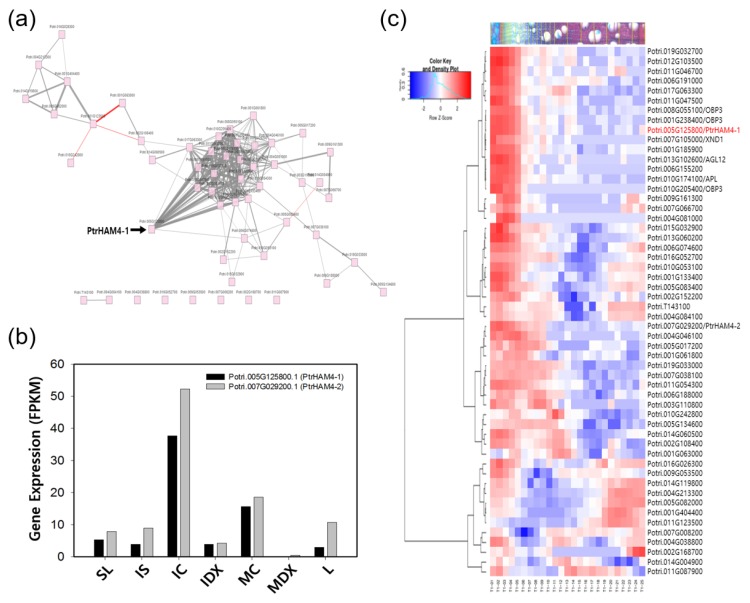
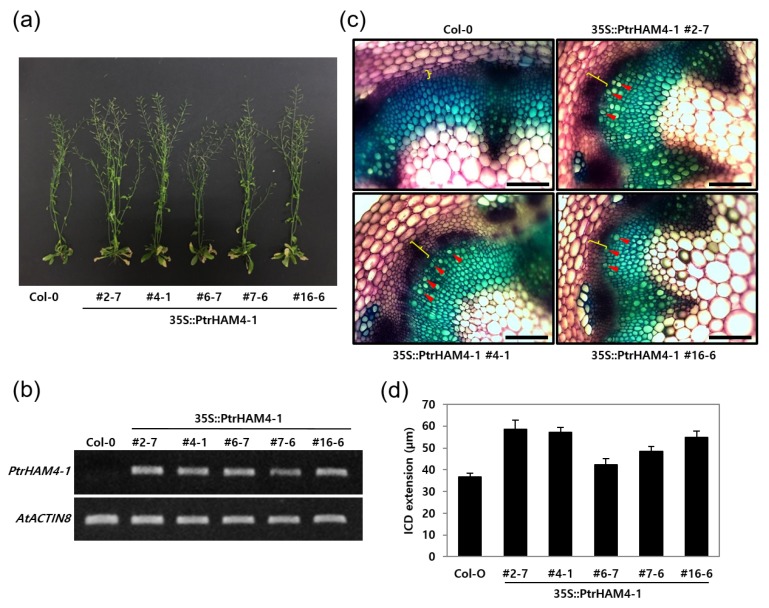
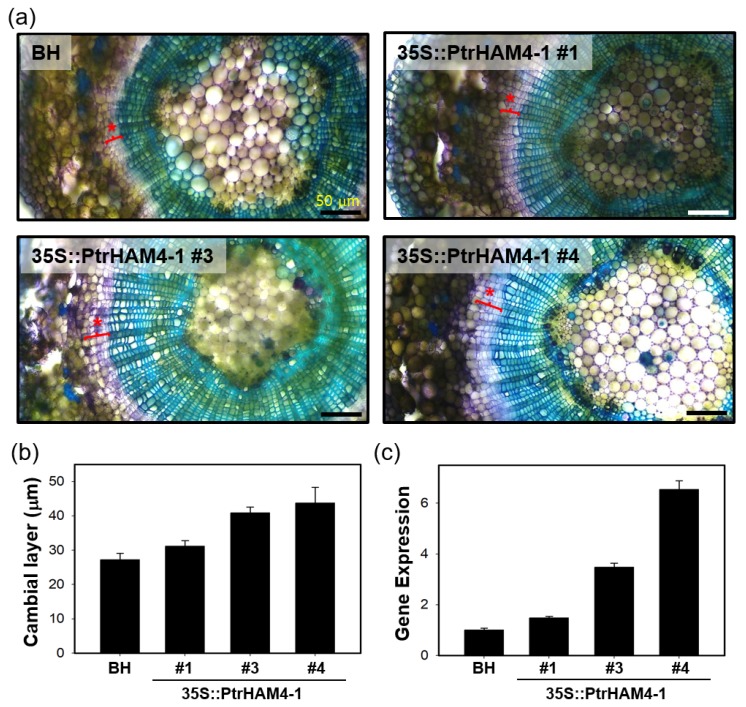
Wood, the most abundant biomass on Earth, is composed of secondary xylem differentiated from vascular cambium. However, the underlying molecular mechanisms of wood formation remain largely unclear. To gain insight into wood formation, we performed a series of wood-forming tissue-specific transcriptome analyses from a hybrid poplar (Populus alba × P. glandulosa, clone BH) using RNA-seq. Together with shoot apex and leaf tissue, cambium and xylem tissues were isolated from vertical stem segments representing a gradient of secondary growth developmental stages (i.e., immature, intermediate, and mature stem). In a comparative transcriptome analysis of the 'developing xylem' and 'leaf' tissue, we could identify critical players catalyzing each biosynthetic step of secondary wall components (e.g., cellulose, xylan, and lignin). Several candidate genes involved in the initiation of vascular cambium formation were found via a co-expression network analysis using abundantly expressed genes in the 'intermediate stem-derived cambium' tissue. We found that transgenic Arabidopsis plants overexpressing the PtrHAM4-1, a GRAS family transcription factor, resulted in a significant increase of vascular cambium development. This phenotype was successfully reproduced in the transgenic poplars overexpressing the PtrHAM4-1. Taken together, our results may serve as a springboard for further research to unravel the molecular mechanism of wood formation, one of the most important biological processes on this planet.
Keywords: Ptrham4-1; poplar; tissue-specific transcriptome; vascular cambium; wood formation.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Downregulating aspen xylan biosynthetic GT43 genes in developing wood stimulates growth via reprograming of the transcriptome.New Phytol. 2018 Jul;219(1):230-245. doi: 10.1111/nph.15160. Epub 2018 Apr 30. New Phytol. 2018. PMID: 29708593
-
WUSCHEL-RELATED HOMEOBOX genes are crucial for normal vascular organization and wood formation in poplar.Plant Sci. 2024 Sep;346:112138. doi: 10.1016/j.plantsci.2024.112138. Epub 2024 May 31. Plant Sci. 2024. PMID: 38825043
-
Transcription factor PagMYB31 positively regulates cambium activity and negatively regulates xylem development in poplar.Plant Cell. 2024 May 1;36(5):1806-1828. doi: 10.1093/plcell/koae040. Plant Cell. 2024. PMID: 38339982 Free PMC article.
-
Genetic and hormonal regulation of cambial development.Physiol Plant. 2013 Jan;147(1):36-45. doi: 10.1111/j.1399-3054.2012.01627.x. Epub 2012 May 3. Physiol Plant. 2013. PMID: 22551327 Review.
-
A review of xylan and lignin biosynthesis: foundation for studying Arabidopsis irregular xylem mutants with pleiotropic phenotypes.Crit Rev Biochem Mol Biol. 2014 May-Jun;49(3):212-41. doi: 10.3109/10409238.2014.889651. Epub 2014 Feb 24. Crit Rev Biochem Mol Biol. 2014. PMID: 24564339 Review.
Cited by
-
Prunus Knotted-like Genes: Genome-Wide Analysis, Transcriptional Response to Cytokinin in Micropropagation, and Rootstock Transformation.Int J Mol Sci. 2023 Feb 3;24(3):3046. doi: 10.3390/ijms24033046. Int J Mol Sci. 2023. PMID: 36769369 Free PMC article.
-
MYB Transcription Factors and Its Regulation in Secondary Cell Wall Formation and Lignin Biosynthesis during Xylem Development.Int J Mol Sci. 2021 Mar 30;22(7):3560. doi: 10.3390/ijms22073560. Int J Mol Sci. 2021. PMID: 33808132 Free PMC article. Review.
-
Genes expression profiles in vascular cambium of Eucalyptus urophylla × Eucalyptus grandis at different ages.BMC Plant Biol. 2023 Oct 18;23(1):500. doi: 10.1186/s12870-023-04500-8. BMC Plant Biol. 2023. PMID: 37848837 Free PMC article.
-
Brassinosteroids affect wood development and properties of Fraxinus mandshurica.Front Plant Sci. 2023 Jul 17;14:1167548. doi: 10.3389/fpls.2023.1167548. eCollection 2023. Front Plant Sci. 2023. PMID: 37546264 Free PMC article.
-
Transcriptome and miRNAs Profiles Reveal Regulatory Network and Key Regulators of Secondary Xylem Formation in "84K" Poplar.Int J Mol Sci. 2023 Nov 17;24(22):16438. doi: 10.3390/ijms242216438. Int J Mol Sci. 2023. PMID: 38003631 Free PMC article.
References
-
- Lal R. Managing soils and ecosystems for mitigating anthropogenic carbon emissions and advancing global food security. BioScience. 2010;60:708–721. doi: 10.1525/bio.2010.60.9.8. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
