

GhFAD2-3 is required for anther development in Gossypium hirsutum
- PMID: 31500565
- PMCID: PMC6734329
- DOI: 10.1186/s12870-019-2010-9
GhFAD2-3 is required for anther development in Gossypium hirsutum
Abstract
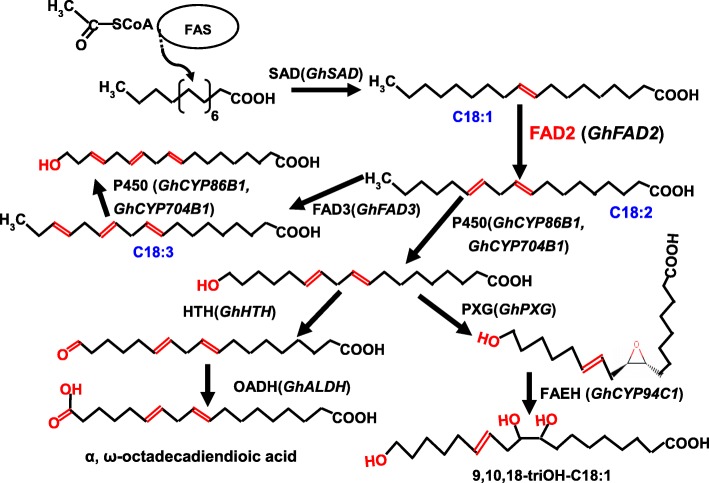
Background: In higher plants, the FAD2 gene encodes the microsomal oleate Δ12-desaturase, one of the key enzymes essential for the biosynthesis of the polyunsaturated lipids that serve many important functions in plant development and stress responses. FAD2 catalyzes the first step, in the biosynthesis of the polyunsaturated fatty acids (PUFAs) found in the cell membrane and cell wall, and it is thus of great importance to investigate the regulatory role of FAD2 in anther development.
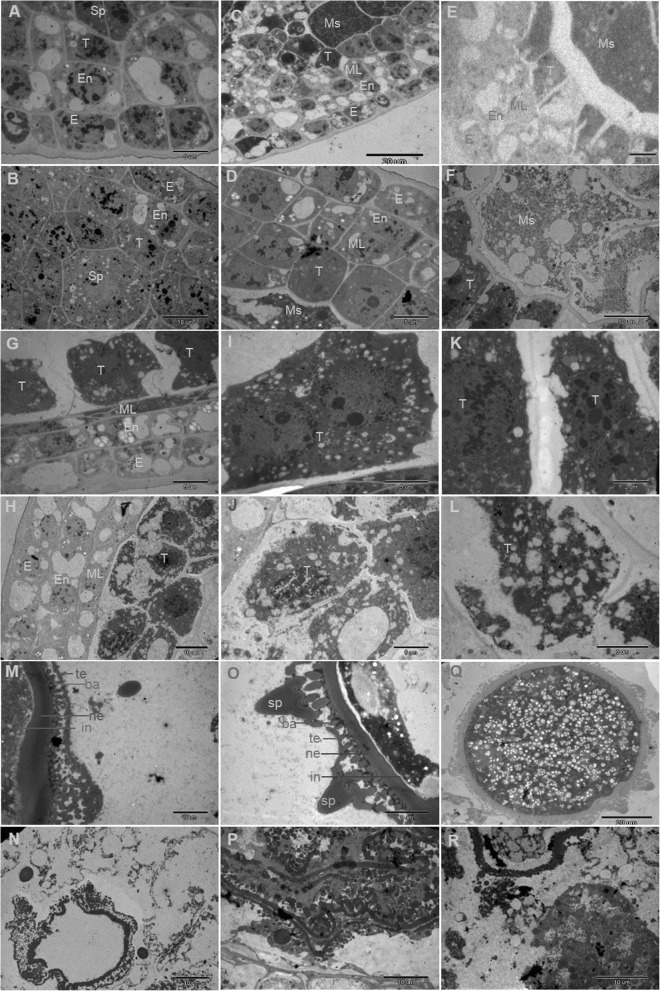
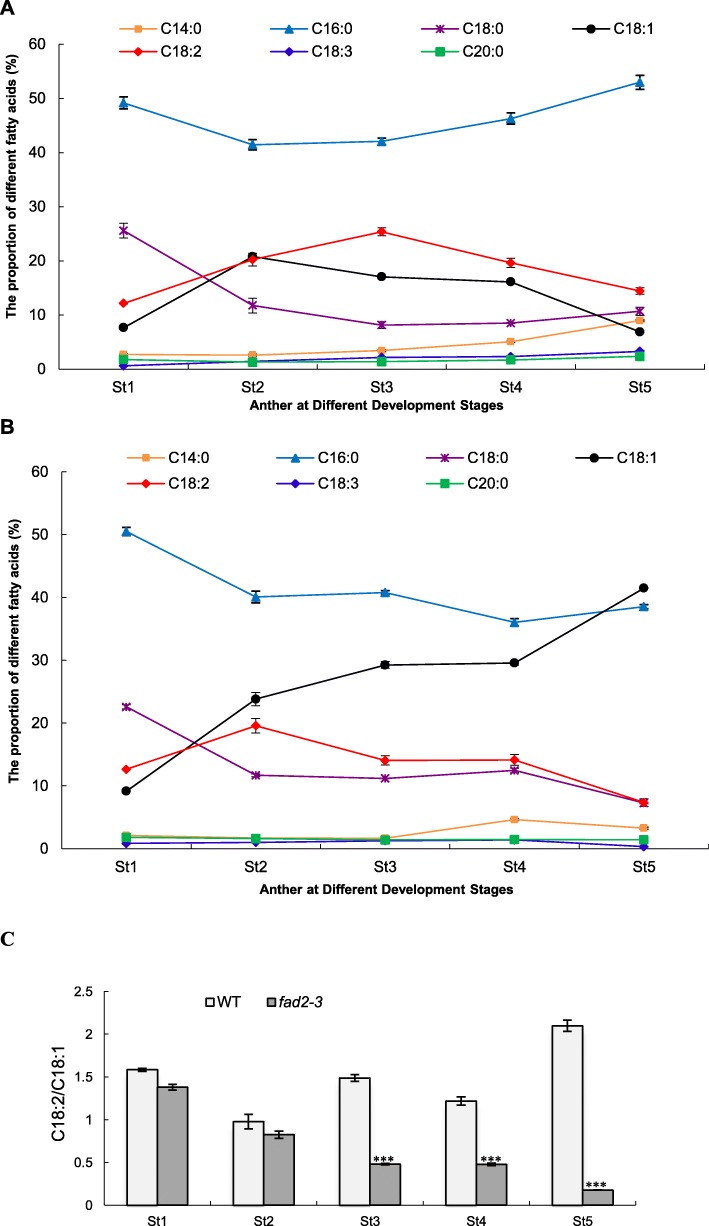
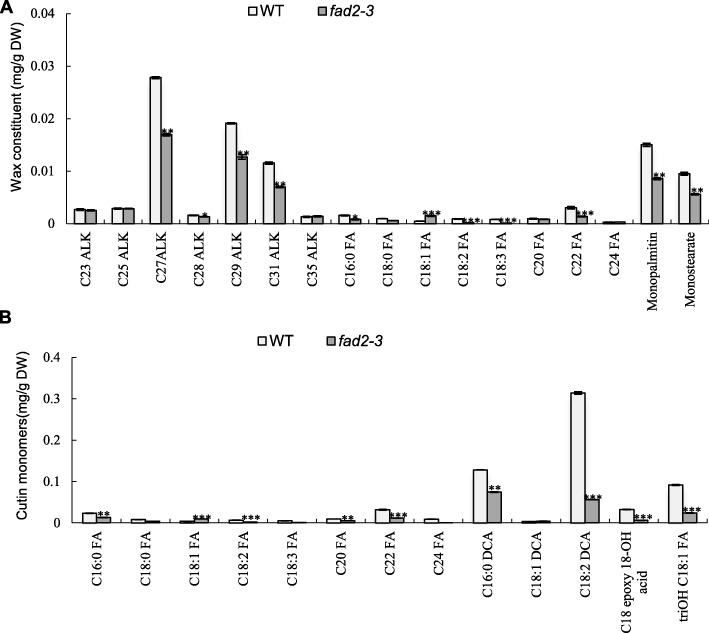
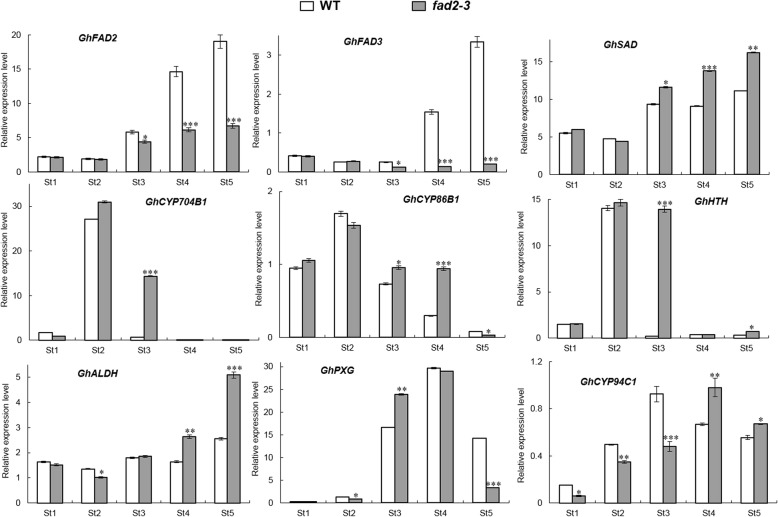
Results: We reported the molecular characterization of the cotton (Gossypium hirsutum) GhFAD2 gene family and the essential role of GhFAD2-3 in cotton anther development. G. hirsutum contains four pairs of homoeologous FAD2 genes (GhFAD2-1 to GhFAD2-4). GhFAD2-3 is ubiquitously and relatively highly expressed in all analyzed tissues, particularly in anthers. Specific inhibition of GhFAD2-3 using the RNA interference approach resulted in male sterility due to impaired anther development at the stages from meiosis to maturation. The cellular phenotypic abnormality observed at the meiosis stage of the GhFAD2-3 silenced plant (fad2-3) coincides with the significant reduction of C18:2 in anthers at the same stage. Compared with that of the wild type (WT), the content of C18:1 was 41.48%, which increased by 5 fold in the fad2-3 anther at the pollen maturation stage. Moreover, the ratio of monounsaturated to polyunsaturated fatty acid was 5.43 in fad2-3 anther, which was much higher than that of the WT (only 0.39). Through compositional analysis of anthers cuticle and transcriptome data, we demonstrated it was unfavorable to the development of anther by regulating GhFAD2-3 expression level to increase the oleic acid content.
Conclusions: Our work demonstrated the importance of C18:2 and/or C18:3 in the development of the pollen exine and anther cuticle in cotton and provided clue for further investigation of the physiological significance of the fatty acid composition for plant growth and development.
Keywords: Fatty acid composition; GhFAD2; Gossypium hirsutum; Linoleic acid; Male sterile; Oleic acid.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



Similar articles
-
Identification and expression of a new delta-12 fatty acid desaturase (FAD2-4) gene in upland cotton and its functional expression in yeast and Arabidopsis thaliana plants.Plant Physiol Biochem. 2009 Jun;47(6):462-71. doi: 10.1016/j.plaphy.2008.12.024. Epub 2009 Jan 18. Plant Physiol Biochem. 2009. PMID: 19217793
-
High-stearic and High-oleic cottonseed oils produced by hairpin RNA-mediated post-transcriptional gene silencing.Plant Physiol. 2002 Aug;129(4):1732-43. doi: 10.1104/pp.001933. Plant Physiol. 2002. PMID: 12177486 Free PMC article.
-
Simultaneous silencing of GhFAD2-1 and GhFATB enhances the quality of cottonseed oil with high oleic acid.J Plant Physiol. 2017 Aug;215:132-139. doi: 10.1016/j.jplph.2017.06.001. Epub 2017 Jun 15. J Plant Physiol. 2017. PMID: 28644971
-
The FAD2 Gene in Plants: Occurrence, Regulation, and Role.Front Plant Sci. 2017 Oct 18;8:1789. doi: 10.3389/fpls.2017.01789. eCollection 2017. Front Plant Sci. 2017. PMID: 29093726 Free PMC article. Review.
-
Structural diversity, biosynthesis, and function of plant falcarin-type polyacetylenic lipids.J Exp Bot. 2022 May 13;73(9):2889-2904. doi: 10.1093/jxb/erac006. J Exp Bot. 2022. PMID: 35560192 Review.
Cited by
-
Explore the gene network regulating the composition of fatty acids in cottonseed.BMC Plant Biol. 2021 Apr 13;21(1):177. doi: 10.1186/s12870-021-02952-4. BMC Plant Biol. 2021. PMID: 33849439 Free PMC article.
-
Insights into the Novel FAD2 Gene Regulating Oleic Acid Accumulation in Peanut Seeds with Different Maturity.Genes (Basel). 2022 Nov 9;13(11):2076. doi: 10.3390/genes13112076. Genes (Basel). 2022. PMID: 36360313 Free PMC article.
-
GhGPAT12/25 Are Essential for the Formation of Anther Cuticle and Pollen Exine in Cotton (Gossypium hirsutum L.).Front Plant Sci. 2021 May 13;12:667739. doi: 10.3389/fpls.2021.667739. eCollection 2021. Front Plant Sci. 2021. PMID: 34054906 Free PMC article.
-
Characterization and transcriptome analysis of a dominant genic male sterile cotton mutant.BMC Plant Biol. 2020 Jul 3;20(1):312. doi: 10.1186/s12870-020-02522-0. BMC Plant Biol. 2020. PMID: 32620078 Free PMC article.
-
Impact of water deficiency on leaf cuticle lipids and gene expression networks in cotton (Gossypium hirsutum L.).BMC Plant Biol. 2022 Aug 17;22(1):404. doi: 10.1186/s12870-022-03788-2. BMC Plant Biol. 2022. PMID: 35978290 Free PMC article.
References
-
- Shanklin John, Cahoon Edgar B. DESATURATION AND RELATED MODIFICATIONS OF FATTY ACIDS. Annual Review of Plant Physiology and Plant Molecular Biology. 1998;49(1):611–641. - PubMed
-
- Jin Un-Ho, Lee Jin-Woo, Chung Young-Soo, Lee Jai-Heon, Yi Young-Byung, Kim Young-Kil, Hyung Nam-In, Pyee Jae-Ho, Chung Chung-Han. Characterization and temporal expression of a ω-6 fatty acid desaturase cDNA from sesame (Sesamum indicum L.) seeds. Plant Science. 2001;161(5):935–941.
-
- Jung Jin Hee, Kim Hyojin, Go Young Sam, Lee Saet Buyl, Hur Cheol-Goo, Kim Hyun Uk, Suh Mi Chung. Identification of functional BrFAD2-1 gene encoding microsomal delta-12 fatty acid desaturase from Brassica rapa and development of Brassica napus containing high oleic acid contents. Plant Cell Reports. 2011;30(10):1881–1892. - PubMed
-
- Li Lingyong, Wang Xiaolin, Gai Junyi, Yu Deyue. Molecular cloning and characterization of a novel microsomal oleate desaturase gene from soybean. Journal of Plant Physiology. 2007;164(11):1516–1526. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
