

Positive Cooperativity in Substrate Binding by Human Thymidylate Synthase
- PMID: 31500803
- PMCID: PMC6818155
- DOI: 10.1016/j.bpj.2019.08.015
Positive Cooperativity in Substrate Binding by Human Thymidylate Synthase
Erratum in
-
Positive Cooperativity in Substrate Binding by Human Thymidylate Synthase.Biophys J. 2021 Sep 21;120(18):4137. doi: 10.1016/j.bpj.2021.08.023. Epub 2021 Aug 24. Biophys J. 2021. PMID: 34433070 Free PMC article. No abstract available.
Abstract
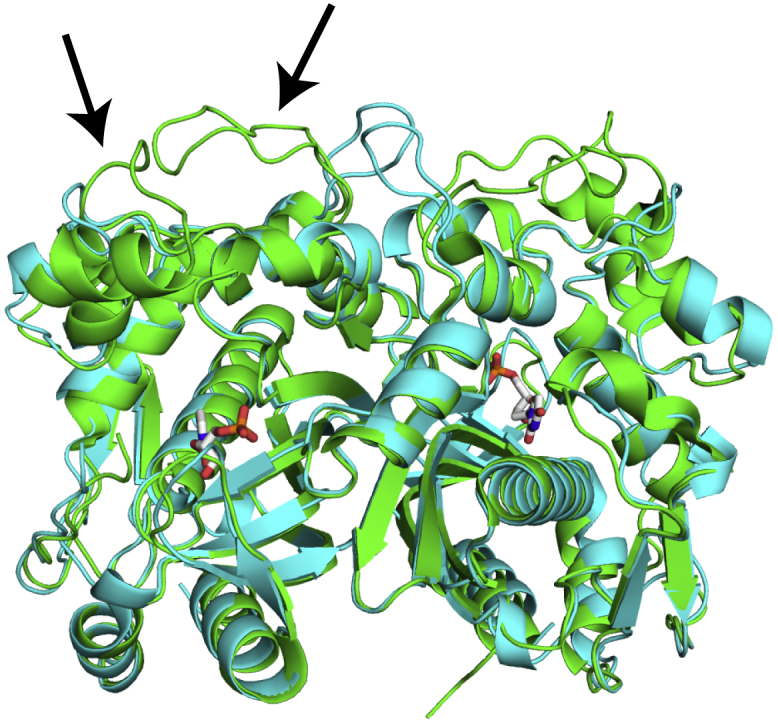
Thymidylate synthase (TS) catalyzes the production of the nucleotide dTMP from deoxyuridine monophosphate (dUMP), making the enzyme necessary for DNA replication and consequently a target for cancer therapeutics. TSs are homodimers with active sites separated by ∼30 Å. Reports of half-the-sites activity in TSs from multiple species demonstrate the presence of allosteric communication between the active sites of this enzyme. A simple explanation for the negative allosteric regulation occurring in half-the-sites activity would be that the two substrates bind with negative cooperativity. However, previous work on Escherichia coli TS revealed that dUMP substrate binds without cooperativity. To gain further insight into TS allosteric function, binding cooperativity in human TS is examined here. Isothermal titration calorimetry and two-dimensional lineshape analysis of NMR titration spectra are used to characterize the thermodynamics of dUMP binding, with a focus on quantification of cooperativity between the two substrate binding events. We find that human TS binds dUMP with ∼9-fold entropically driven positive cooperativity (ρITC = 9 ± 1, ρNMR = 7 ± 1), in contrast to the apparent strong negative cooperativity reported previously. Our work further demonstrates the necessity of globally fitting isotherms collected under various conditions, as well as accurate determination of binding competent protein concentration, for calorimetric characterization of homotropic cooperative binding. Notably, an initial curvature of the isotherm is found to be indicative of positively cooperative binding. Two-dimensional lineshape analysis NMR is also found to be an informative tool for quantifying binding cooperativity, particularly in cases in which bound intermediates yield unique resonances.
Copyright © 2019 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Monod J., Wyman J., Changeux J.P. On the nature of allosteric transitions: a plausible model. J. Mol. Biol. 1965;12:88–118. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
