

Chromosomal Recombination Targets in Chlamydia Interspecies Lateral Gene Transfer
- PMID: 31501285
- PMCID: PMC6832074
- DOI: 10.1128/JB.00365-19
Chromosomal Recombination Targets in Chlamydia Interspecies Lateral Gene Transfer
Abstract
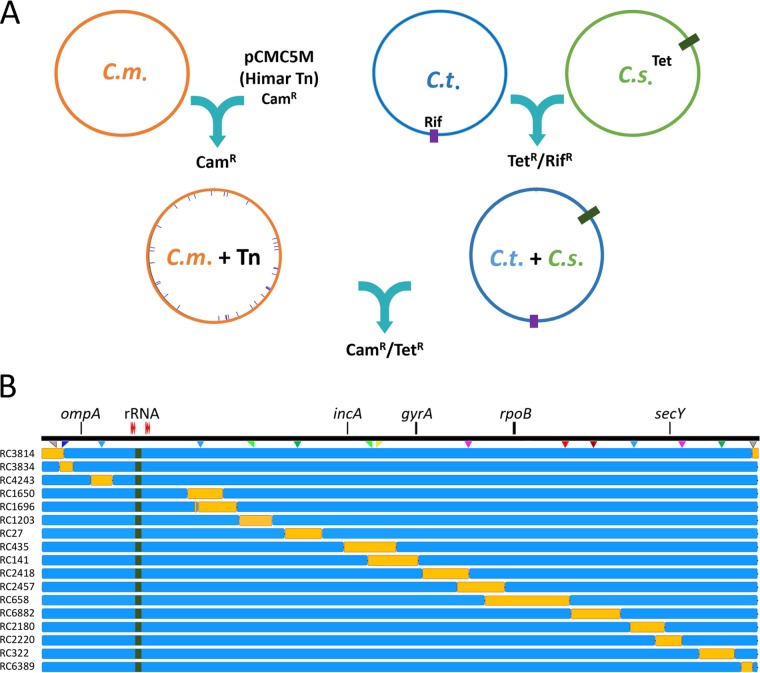
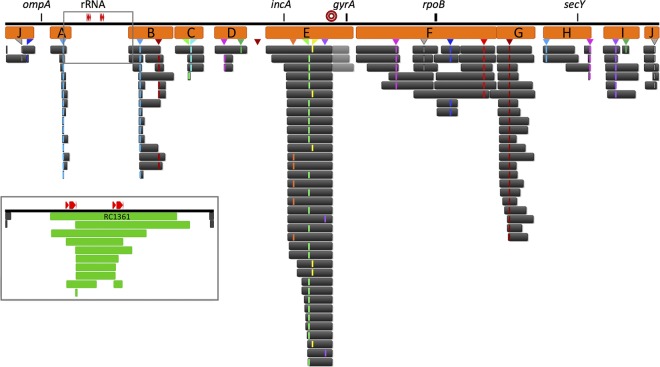
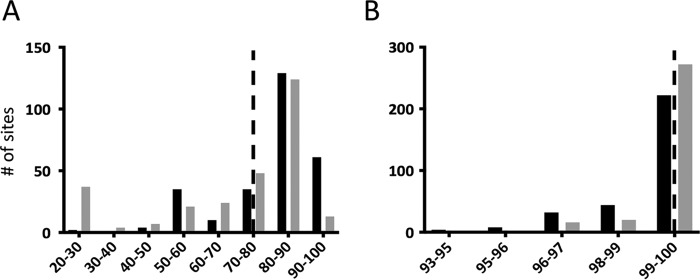
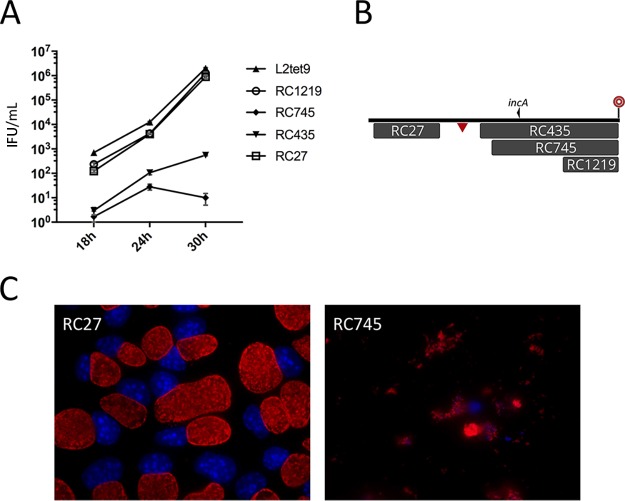
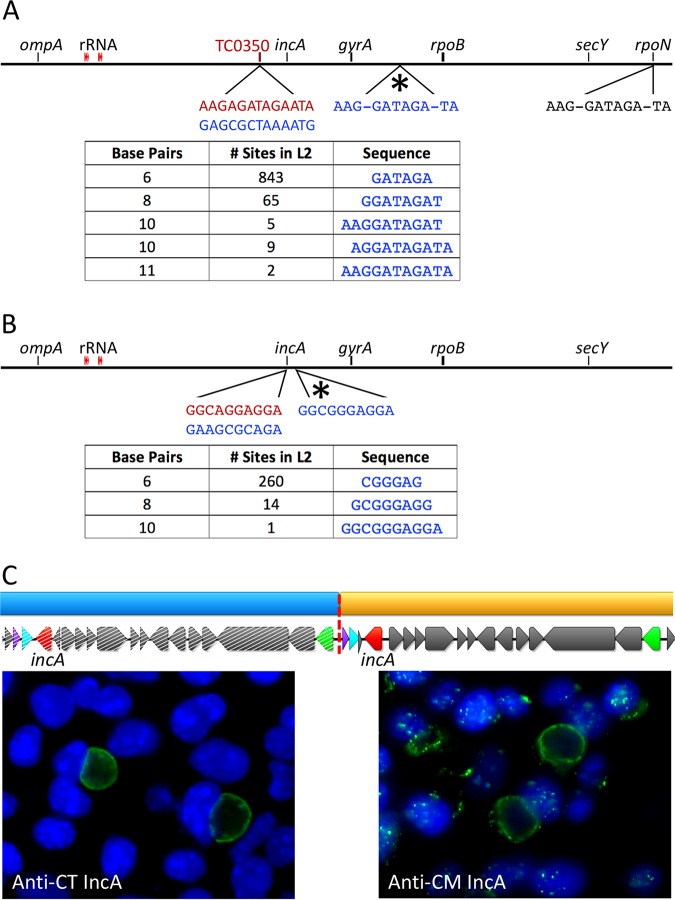
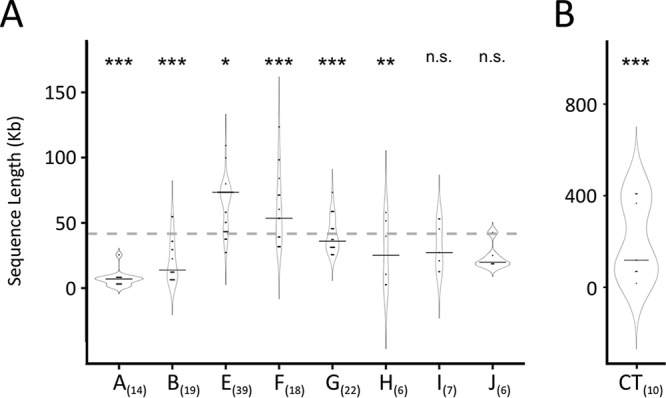
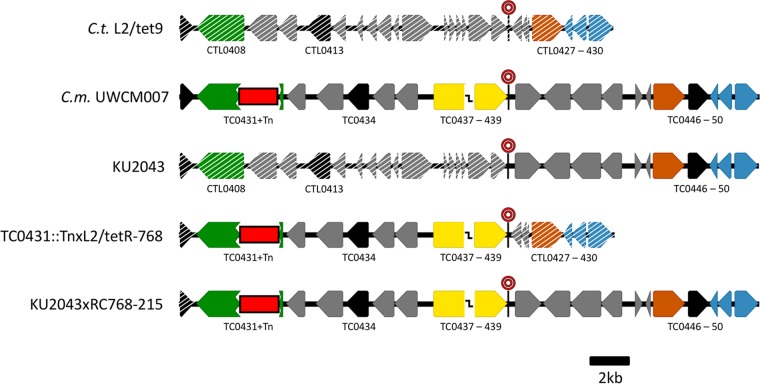
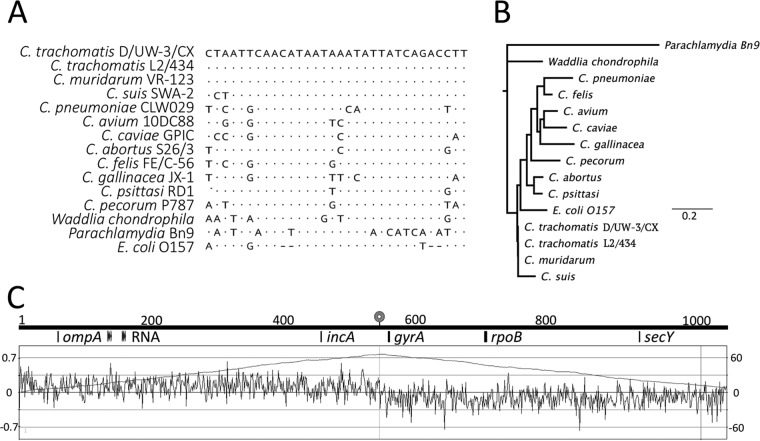
Lateral gene transfer (LGT) among Chlamydia trachomatis strains is common, in both isolates generated in the laboratory and those examined directly from patients. In contrast, there are very few examples of recent acquisition of DNA by any Chlamydia spp. from any other species. Interspecies LGT in this system was analyzed using crosses of tetracycline (Tc)-resistant C. trachomatis L2/434 and chloramphenicol (Cam)-resistant C. muridarum VR-123. Parental C. muridarum strains were created using a plasmid-based Himar transposition system, which led to integration of the Camr marker randomly across the chromosome. Fragments encompassing 79% of the C. muridarum chromosome were introduced into a C. trachomatis background, with the total coverage contained on 142 independent recombinant clones. Genome sequence analysis of progeny strains identified candidate recombination hot spots, a property not consistent with in vitroC. trachomatis × C. trachomatis (intraspecies) crosses. In both interspecies and intraspecies crosses, there were examples of duplications, mosaic recombination endpoints, and recombined sequences that were not linked to the selection marker. Quantitative analysis of the distribution and constitution of inserted sequences indicated that there are different constraints on interspecies LGT than on intraspecies crosses. These constraints may help explain why there is so little evidence of interspecies genetic exchange in this system, which is in contrast to very widespread intraspecies exchange in C. trachomatisIMPORTANCE Genome sequence analysis has demonstrated that there is widespread lateral gene transfer among strains within the species C. trachomatis and with other closely related Chlamydia species in laboratory experiments. This is in contrast to the complete absence of foreign DNA in the genomes of sequenced clinical C. trachomatis strains. There is no understanding of any mechanisms of genetic transfer in this important group of pathogens. In this report, we demonstrate that interspecies genetic exchange can occur but that the nature of the fragments exchanged is different than those observed in intraspecies crosses. We also generated a large hybrid strain library that can be exploited to examine important aspects of chlamydial disease.
Keywords: Chlamydia; lateral gene transfer; recombination; transposon.
Copyright © 2019 American Society for Microbiology.
Figures
References
-
- Caldwell HD, Wood H, Crane D, Bailey R, Jones RB, Mabey D, Maclean I, Mohammed Z, Peeling R, Roshick C, Schachter J, Solomon AW, Stamm WE, Suchland RJ, Taylor L, West SK, Quinn TC, Belland RJ, McClarty G. 2003. Polymorphisms in Chlamydia trachomatis tryptophan synthase genes differentiate between genital and ocular isolates. J Clin Invest 111:1757–1769. doi:10.1172/JCI17993. - DOI - PMC - PubMed
-
- Read TD, Brunham RC, Shen C, Gill SR, Heidelberg JF, White O, Hickey EK, Peterson J, Utterback T, Berry K, Bass S, Linher K, Weidman J, Khouri H, Craven B, Bowman C, Dodson R, Gwinn M, Nelson W, DeBoy R, Kolonay J, McClarty G, Salzberg SL, Eisen J, Fraser CM. 2000. Genome sequences of Chlamydia trachomatis MoPn and Chlamydia pneumoniae AR39. Nucleic Acids Res 28:1397–1406. doi:10.1093/nar/28.6.1397. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
