

Distinct intraspecies virulence mechanisms regulated by a conserved transcription factor
- PMID: 31501343
- PMCID: PMC6765310
- DOI: 10.1073/pnas.1903461116
Distinct intraspecies virulence mechanisms regulated by a conserved transcription factor
Abstract
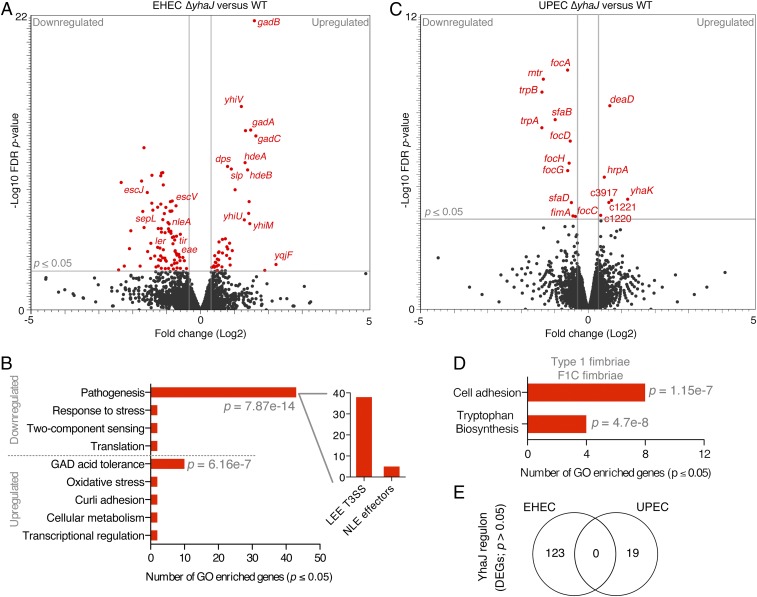
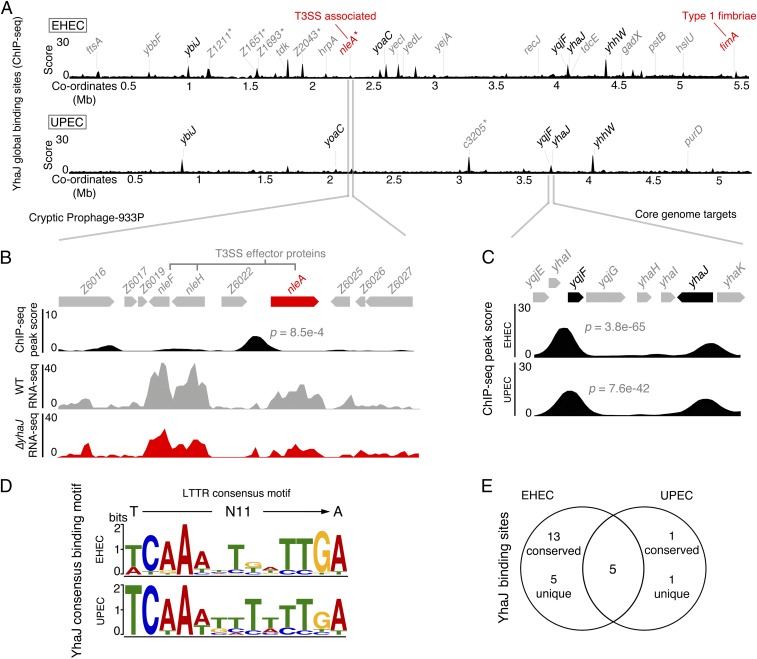
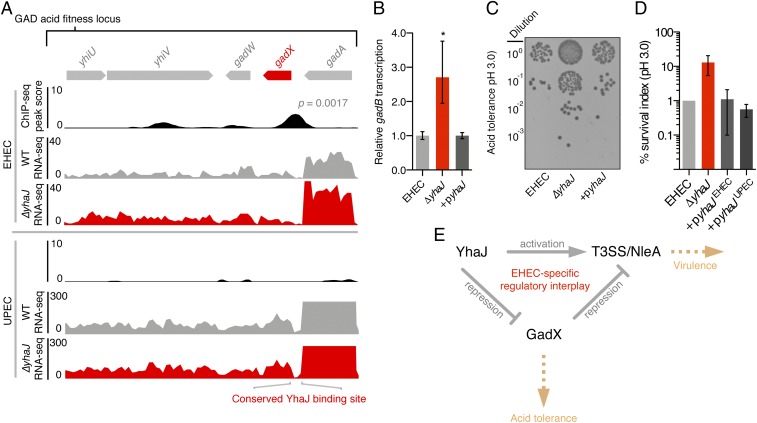
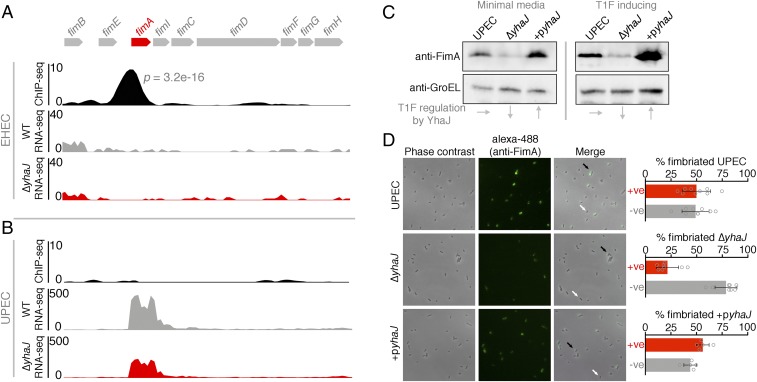
Tailoring transcriptional regulation to coordinate the expression of virulence factors in tandem with the core genome is a hallmark of bacterial pathogen evolution. Bacteria encode hundreds of transcription factors forming the base-level control of gene regulation. Moreover, highly homologous regulators are assumed to control conserved genes between members within a species that harbor the same genetic targets. We have explored this concept in 2 Escherichia coli pathotypes that employ distinct virulence mechanisms that facilitate specification of a different niche within the host. Strikingly, we found that the transcription factor YhaJ actively regulated unique gene sets between intestinal enterohemorrhagic E. coli (EHEC) and extraintestinal uropathogenic E. coli (UPEC), despite being very highly conserved. In EHEC, YhaJ directly activates expression of type 3 secretion system components and effectors. Alternatively, YhaJ enhances UPEC virulence regulation by binding directly to the phase-variable type 1 fimbria promoter, driving its expression. Additionally, YhaJ was found to override the universal GAD acid tolerance system but exclusively in EHEC, thereby indirectly enhancing type 3 secretion pleiotropically. These results have revealed that within a species, conserved regulators are actively repurposed in a "personalized" manner to benefit particular lifestyles and drive virulence via multiple distinct mechanisms.
Keywords: gene expression; niche; regulation; type 1 fimbriae; type 3 secretion.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
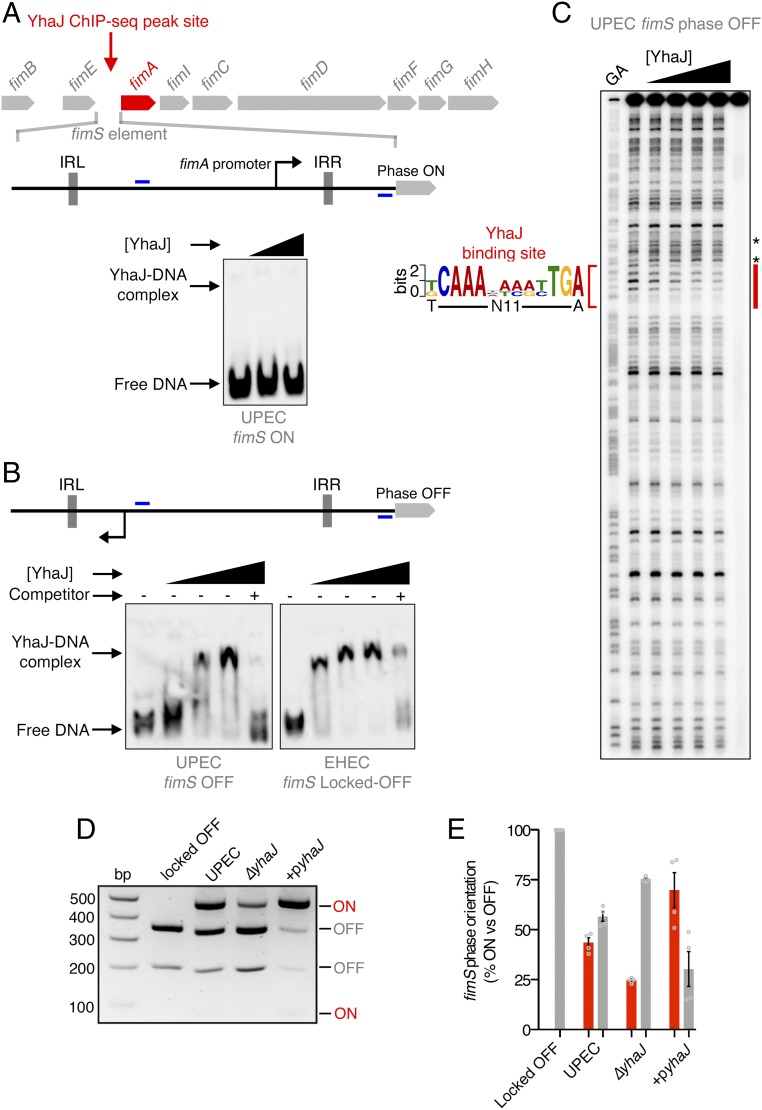
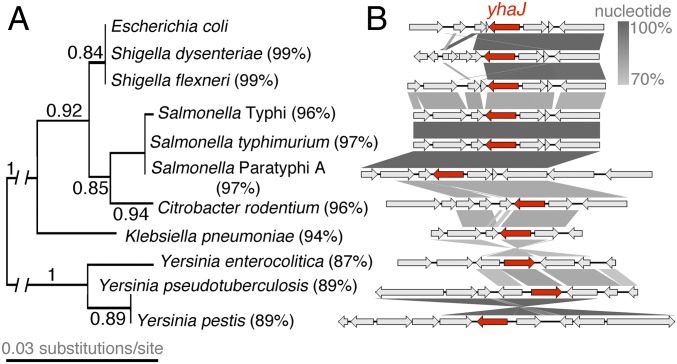
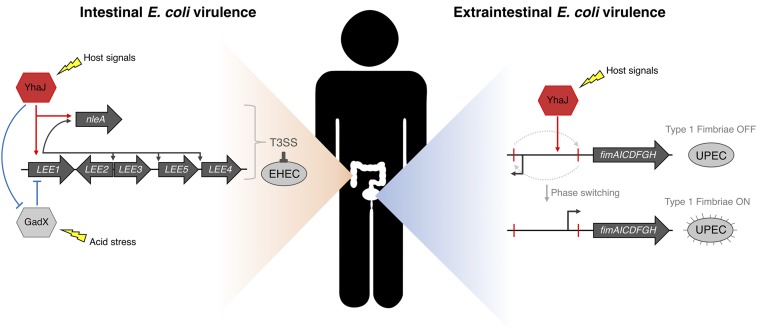
References
-
- Browning D. F., Busby S. J. W., Local and global regulation of transcription initiation in bacteria. Nat. Rev. Microbiol. 14, 638–650 (2016). - PubMed
-
- Seshasayee A. S. N., Sivaraman K., Luscombe N. M., “An overview of prokaryotic transcription factors” in A Handbook of Transcription Factors, Hughes T., Ed. (Springer, 2011), vol. 52, pp. 7–23. - PubMed
-
- Moran N. A., Microbial minimalism: Genome reduction in bacterial pathogens. Cell 108, 583–586 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
