

Nucleosome Dynamics: a new tool for the dynamic analysis of nucleosome positioning
- PMID: 31504766
- PMCID: PMC6765203
- DOI: 10.1093/nar/gkz759
Nucleosome Dynamics: a new tool for the dynamic analysis of nucleosome positioning
Abstract
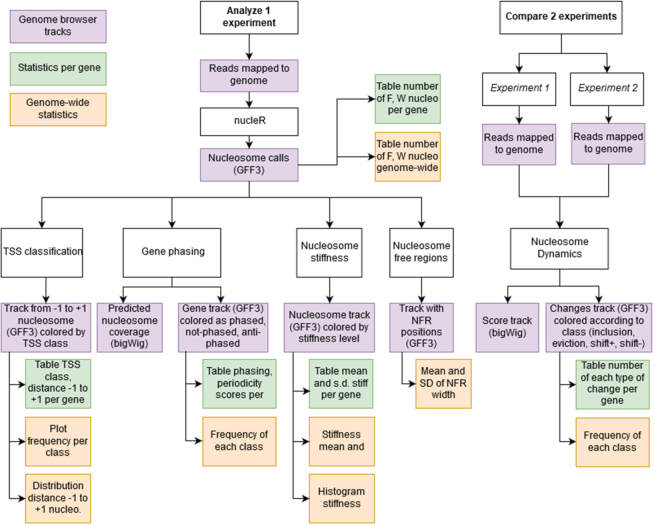
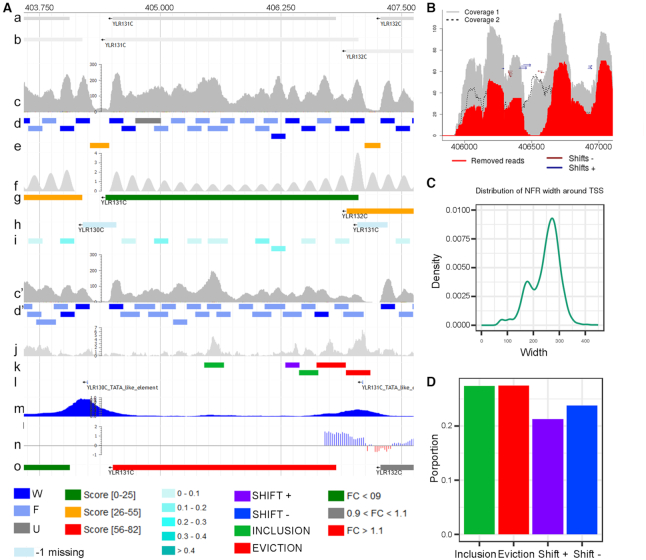
We present Nucleosome Dynamics, a suite of programs integrated into a virtual research environment and created to define nucleosome architecture and dynamics from noisy experimental data. The package allows both the definition of nucleosome architectures and the detection of changes in nucleosomal organization due to changes in cellular conditions. Results are displayed in the context of genomic information thanks to different visualizers and browsers, allowing the user a holistic, multidimensional view of the genome/transcriptome. The package shows good performance for both locating equilibrium nucleosome architecture and nucleosome dynamics and provides abundant useful information in several test cases, where experimental data on nucleosome position (and for some cases expression level) have been collected for cells under different external conditions (cell cycle phase, yeast metabolic cycle progression, changes in nutrients or difference in MNase digestion level). Nucleosome Dynamics is a free software and is provided under several distribution models.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures




Similar articles
-
Nucleosome positioning and nucleosome stacking: two faces of the same coin.Mol Biosyst. 2012 Apr;8(4):1172-8. doi: 10.1039/c2mb05407h. Epub 2012 Jan 23. Mol Biosyst. 2012. PMID: 22266567
-
Z curve theory-based analysis of the dynamic nature of nucleosome positioning in Saccharomyces cerevisiae.Gene. 2013 Nov 1;530(1):8-18. doi: 10.1016/j.gene.2013.08.018. Epub 2013 Aug 16. Gene. 2013. PMID: 23958656
-
Sub-nucleosomal Genome Structure Reveals Distinct Nucleosome Folding Motifs.Cell. 2019 Jan 24;176(3):520-534.e25. doi: 10.1016/j.cell.2018.12.014. Epub 2019 Jan 17. Cell. 2019. PMID: 30661750
-
Establishing nucleosome architecture and stability at promoters: Roles of pioneer transcription factors and the RSC chromatin remodeler.Bioessays. 2017 May;39(5). doi: 10.1002/bies.201600237. Epub 2017 Mar 27. Bioessays. 2017. PMID: 28345796 Review.
-
Nucleosome positioning in yeasts: methods, maps, and mechanisms.Chromosoma. 2015 Jun;124(2):131-51. doi: 10.1007/s00412-014-0501-x. Epub 2014 Dec 23. Chromosoma. 2015. PMID: 25529773 Review.
Cited by
-
MiOS, an integrated imaging and computational strategy to model gene folding with nucleosome resolution.Nat Struct Mol Biol. 2022 Oct;29(10):1011-1023. doi: 10.1038/s41594-022-00839-y. Epub 2022 Oct 11. Nat Struct Mol Biol. 2022. PMID: 36220894 Free PMC article.
-
Inferring differential protein binding from time-series chromatin accessibility data.Bioinform Adv. 2025 Apr 10;5(1):vbaf080. doi: 10.1093/bioadv/vbaf080. eCollection 2025. Bioinform Adv. 2025. PMID: 40297777 Free PMC article.
-
Genome-wide chromatin interaction map for Trypanosoma cruzi.Nat Microbiol. 2023 Nov;8(11):2103-2114. doi: 10.1038/s41564-023-01483-y. Epub 2023 Oct 12. Nat Microbiol. 2023. PMID: 37828247 Free PMC article.
-
Prohibitin 2 orchestrates long noncoding RNA and gene transcription to accelerate tumorigenesis.Nat Commun. 2024 Sep 27;15(1):8385. doi: 10.1038/s41467-024-52425-z. Nat Commun. 2024. PMID: 39333493 Free PMC article.
-
The PCNA-Pol δ complex couples lagging strand DNA synthesis to parental histone transfer for epigenetic inheritance.Sci Adv. 2024 Jun 7;10(23):eadn5175. doi: 10.1126/sciadv.adn5175. Epub 2024 Jun 5. Sci Adv. 2024. PMID: 38838138 Free PMC article.
References
-
- Barski A., Cuddapah S., Cui K., Roh T.-Y., Schones D.E., Wang Z., Wei G., Chepelev I., Zhao K.. High-Resolution profiling of histone methylations in the human genome. Cell. 2007; 129:823–837. - PubMed
-
- Yuan G.-C. Genome-Scale Identification of Nucleosome Positions in S. cerevisiae. Science. 2005; 309:626–630. - PubMed
-
- Rando O.J., Ahmad K.. Rules and regulation in the primary structure of chromatin. Curr. Opin. Cell Biol. 2007; 19:250–256. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
