

Simultaneous targeting of DNA replication and homologous recombination in glioblastoma with a polyether ionophore
- PMID: 31504812
- PMCID: PMC7442340
- DOI: 10.1093/neuonc/noz159
Simultaneous targeting of DNA replication and homologous recombination in glioblastoma with a polyether ionophore
Abstract
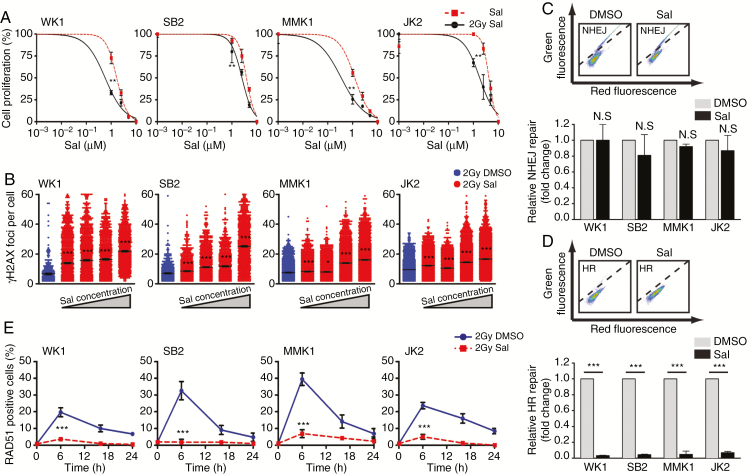
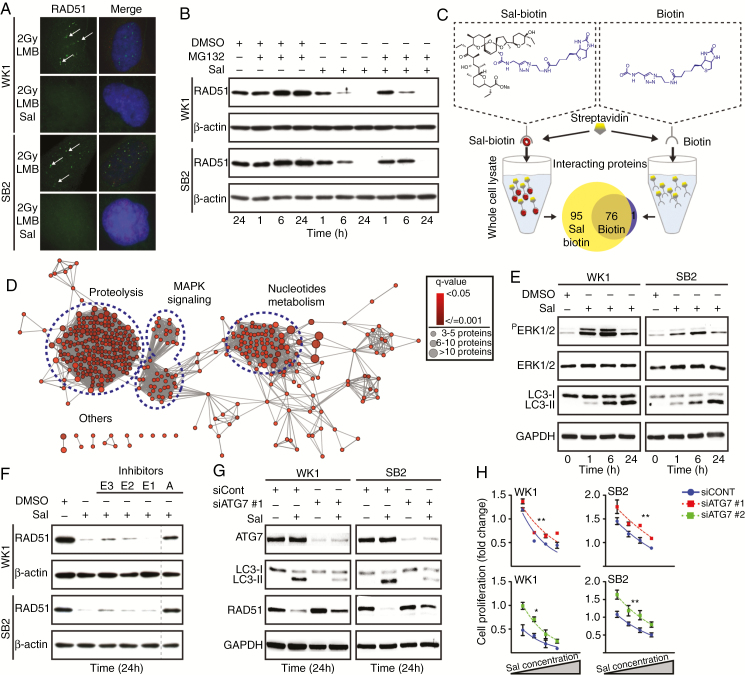
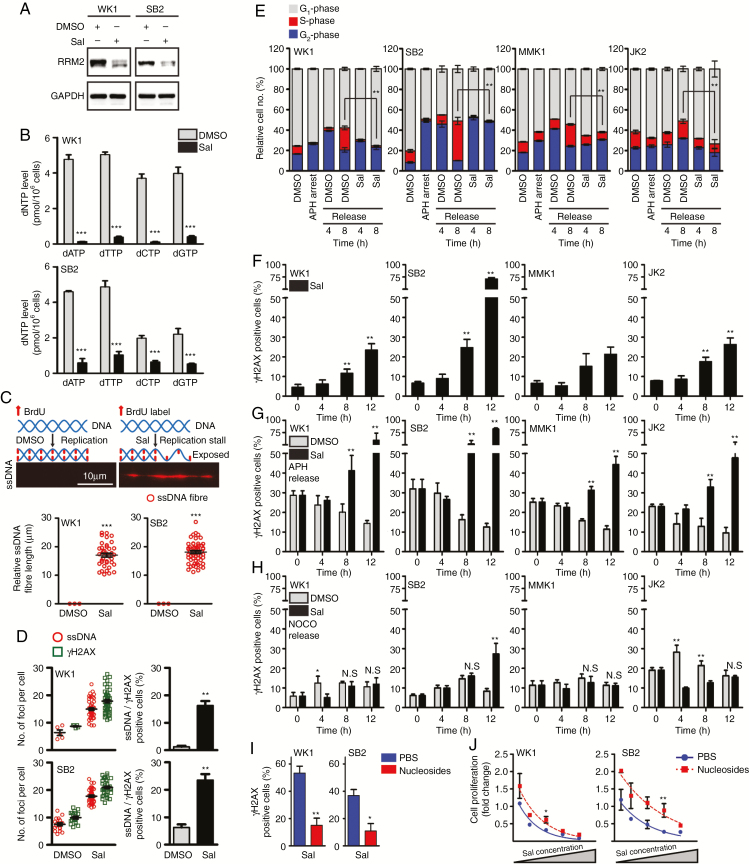
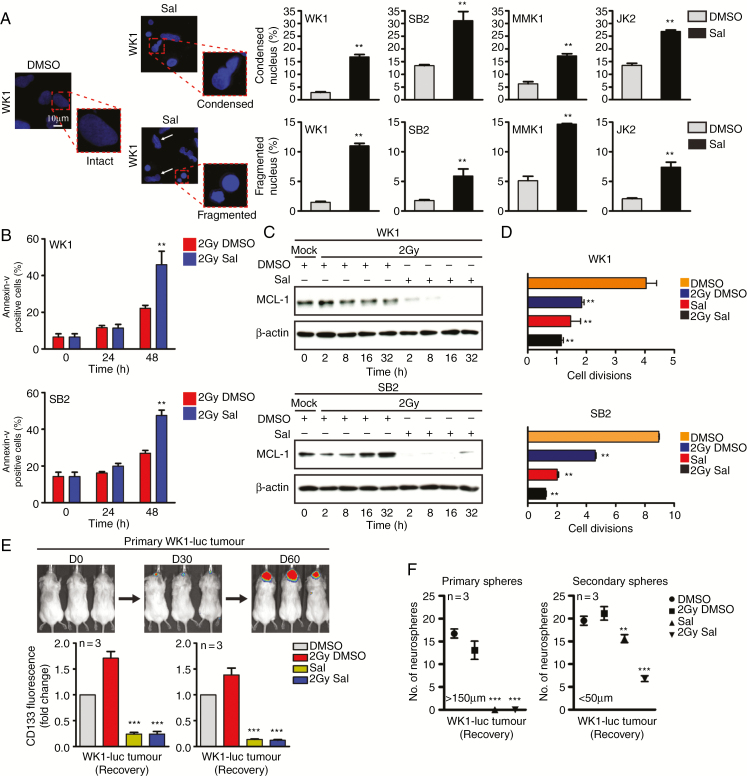
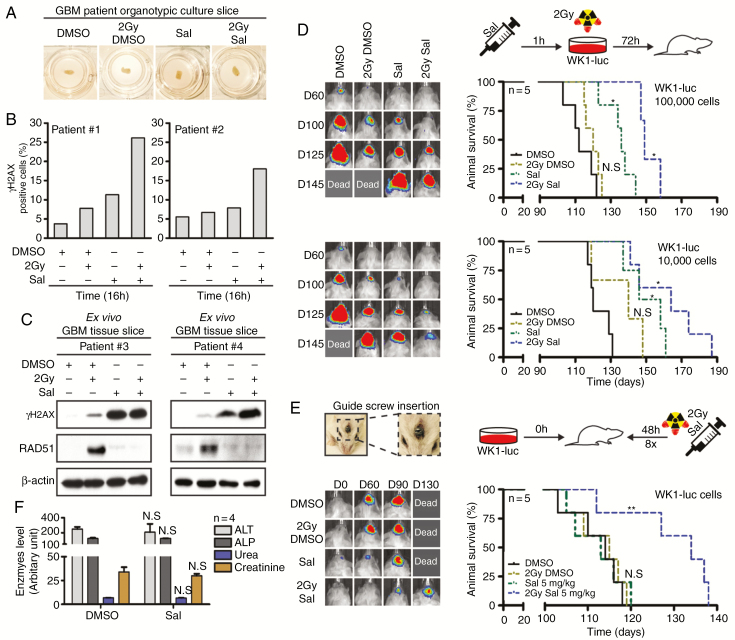
Background: Despite significant endeavor having been applied to identify effective therapies to treat glioblastoma (GBM), survival outcomes remain intractable. The greatest nonsurgical benefit arises from radiotherapy, though tumors typically recur due to robust DNA repair. Patients could therefore benefit from therapies with the potential to prevent DNA repair and synergize with radiotherapy. In this work, we investigated the potential of salinomycin to enhance radiotherapy and further uncover novel dual functions of this ionophore to induce DNA damage and prevent repair.
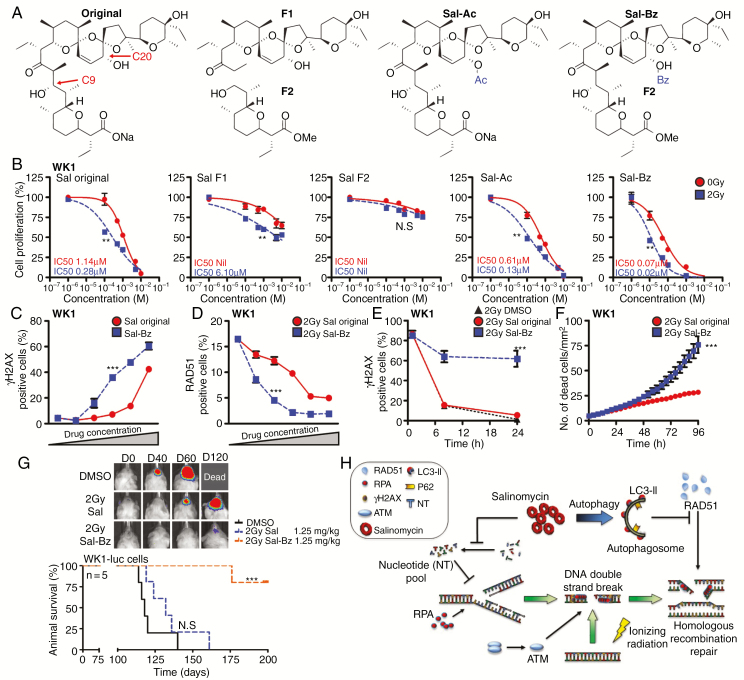
Methods: In vitro primary GBM models and ex vivo GBM patient explants were used to determine the mechanism of action of salinomycin by immunoblot, flow cytometry, immunofluorescence, immunohistochemistry, and mass spectrometry. In vivo efficacy studies were performed using orthotopic GBM animal xenograft models. Salinomycin derivatives were synthesized to increase drug efficacy and explore structure-activity relationships.
Results: Here we report novel dual functions of salinomycin. Salinomycin induces toxic DNA lesions and prevents subsequent recovery by targeting homologous recombination (HR) repair. Salinomycin appears to target the more radioresistant GBM stem cell-like population and synergizes with radiotherapy to significantly delay tumor formation in vivo. We further developed salinomycin derivatives which display greater efficacy in vivo while retaining the same beneficial mechanisms of action.
Conclusion: Our findings highlight the potential of salinomycin to induce DNA lesions and inhibit HR to greatly enhance the effect of radiotherapy. Importantly, first-generation salinomycin derivatives display greater efficacy and may pave the way for clinical testing of these agents.
Keywords: DNA damage; drug discovery; glioblastoma; homologous recombination.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Neuro-Oncology.
Figures
Comment in
-
Salinomycin targets the genome of radioresistant cells in glioblastomas.Neuro Oncol. 2020 Feb 20;22(2):167-168. doi: 10.1093/neuonc/noz224. Neuro Oncol. 2020. PMID: 31784742 Free PMC article. No abstract available.
References
-
- Hegi ME, Diserens AC, Gorlia T, et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N Engl J Med. 2005;352(10):997–1003. - PubMed
-
- Bao S, Wu Q, McLendon RE, et al. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature. 2006;444(7120):756–760. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
