

Surface loops of trypsin-like serine proteases as determinants of function
- PMID: 31505212
- PMCID: PMC7615277
- DOI: 10.1016/j.biochi.2019.09.004
Surface loops of trypsin-like serine proteases as determinants of function
Abstract
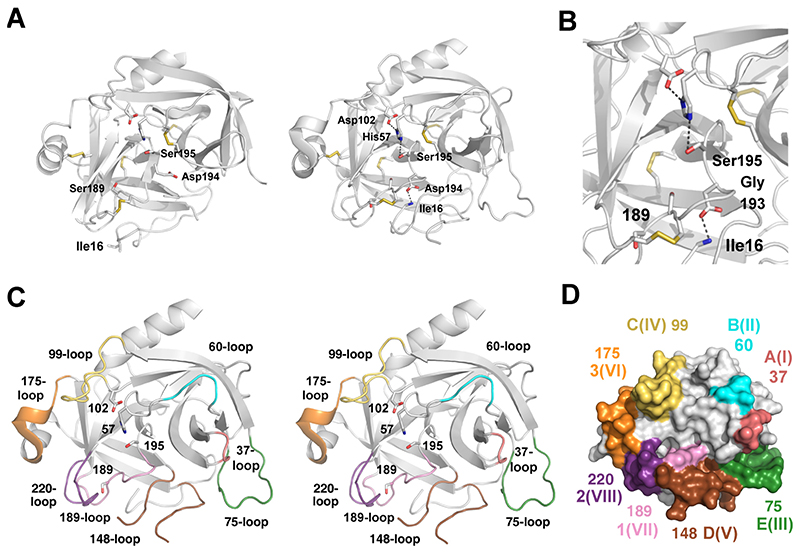
Trypsin and chymotrypsin-like serine proteases from family S1 (clan PA) constitute the largest protease group in humans and more generally in vertebrates. The prototypes chymotrypsin, trypsin and elastase represent simple digestive proteases in the gut, where they cleave nearly any protein. Multidomain trypsin-like proteases are key players in the tightly controlled blood coagulation and complement systems, as well as related proteases that are secreted from diverse immune cells. Some serine proteases are expressed in nearly all tissues and fluids of the human body, such as the human kallikreins and kallikrein-related peptidases with specialization for often unique substrates and accurate timing of activity. HtrA and membrane-anchored serine proteases fulfill important physiological tasks with emerging roles in cancer. The high diversity of all family members, which share the tandem β-barrel architecture of the chymotrypsin-fold in the catalytic domain, is conferred by the large differences of eight surface loops, surrounding the active site. The length of these loops alters with insertions and deletions, resulting in remarkably different three-dimensional arrangements. In addition, metal binding sites for Na+, Ca2+ and Zn2+ serve as regulatory elements, as do N-glycosylation sites. Depending on the individual tasks of the protease, the surface loops determine substrate specificity, control the turnover and allow regulation of activation, activity and degradation by other proteins, which are often serine proteases themselves. Most intriguingly, in some serine proteases, the surface loops interact as allosteric network, partially tuned by protein co-factors. Knowledge of these subtle and complicated molecular motions may allow nowadays for new and specific pharmaceutical or medical approaches.
Keywords: Allosteric network; Chymotrypsin fold; Regulatory mechanism; Structure-function relationship; Surface loops; Zymogenicity.
Copyright © 2019 Elsevier B.V. and Société Française de Biochimie et Biologie Moléculaire (SFBBM). All rights reserved.
Figures







Similar articles
-
The structure of MBL-associated serine protease-2 reveals that identical substrate specificities of C1s and MASP-2 are realized through different sets of enzyme-substrate interactions.J Mol Biol. 2004 Oct 1;342(5):1533-46. doi: 10.1016/j.jmb.2004.07.014. J Mol Biol. 2004. PMID: 15364579
-
Residue 225 determines the Na(+)-induced allosteric regulation of catalytic activity in serine proteases.Proc Natl Acad Sci U S A. 1996 Oct 1;93(20):10653-6. doi: 10.1073/pnas.93.20.10653. Proc Natl Acad Sci U S A. 1996. PMID: 8855234 Free PMC article.
-
Modeling and structural analysis of PA clan serine proteases.BMC Res Notes. 2012 May 24;5:256. doi: 10.1186/1756-0500-5-256. BMC Res Notes. 2012. PMID: 22624962 Free PMC article.
-
Determinants of specificity in coagulation proteases.J Thromb Haemost. 2005 Nov;3(11):2401-8. doi: 10.1111/j.1538-7836.2005.01456.x. J Thromb Haemost. 2005. PMID: 16241939 Review.
-
Membrane anchored serine proteases: a rapidly expanding group of cell surface proteolytic enzymes with potential roles in cancer.Cancer Metastasis Rev. 2003 Jun-Sep;22(2-3):237-58. doi: 10.1023/a:1023003616848. Cancer Metastasis Rev. 2003. PMID: 12784999 Review.
Cited by
-
FAM111A regulates replication origin activation and cell fitness.Life Sci Alliance. 2023 Oct 4;6(12):e202302111. doi: 10.26508/lsa.202302111. Print 2023 Dec. Life Sci Alliance. 2023. PMID: 37793778 Free PMC article.
-
The structure of the Gemella haemolysans M26 IgA1 protease trypsin-like domain.Acta Crystallogr F Struct Biol Commun. 2025 Apr 1;81(Pt 4):124-129. doi: 10.1107/S2053230X25001219. Epub 2025 Feb 28. Acta Crystallogr F Struct Biol Commun. 2025. PMID: 40019192 Free PMC article.
-
A conserved Phytophthora apoplastic trypsin-like serine protease targets the receptor-like kinase BAK1 to dampen plant immunity.Nat Plants. 2025 Jul;11(7):1401-1415. doi: 10.1038/s41477-025-02039-0. Epub 2025 Jul 1. Nat Plants. 2025. PMID: 40596719
-
Probing activation-driven changes in coagulation factor IX by mass spectrometry.J Thromb Haemost. 2021 Jun;19(6):1447-1459. doi: 10.1111/jth.15288. Epub 2021 Apr 5. J Thromb Haemost. 2021. PMID: 33687765 Free PMC article.
-
Structure-mechanics statistical learning uncovers mechanical relay in proteins.Chem Sci. 2022 Jan 19;13(13):3688-3696. doi: 10.1039/d1sc06184d. eCollection 2022 Mar 30. Chem Sci. 2022. PMID: 35432911 Free PMC article.
References
-
- Matthews BW, Sigler PB, Henderson R, Blow DM. Three-dimensional Structure of Tosyl-α-chymotrypsin. Nature. 1967;214:652–656. - PubMed
-
- Hartley BS, Neurath H. Homologies in Serine Proteinases [and Discussion] Phil Trans R Soc B. 1970;257:77–87. - PubMed
-
- Gorbalenya AE, Donchenko AP, Blinov VM, Koonin EV. Cysteine proteases of positive strand RNA viruses and chymotrypsin-like serine proteases. Febs Lett. 1989;243:103–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
