

ATF6 regulates the development of chronic pancreatitis by inducing p53-mediated apoptosis
- PMID: 31506423
- PMCID: PMC6737032
- DOI: 10.1038/s41419-019-1919-0
ATF6 regulates the development of chronic pancreatitis by inducing p53-mediated apoptosis
Abstract
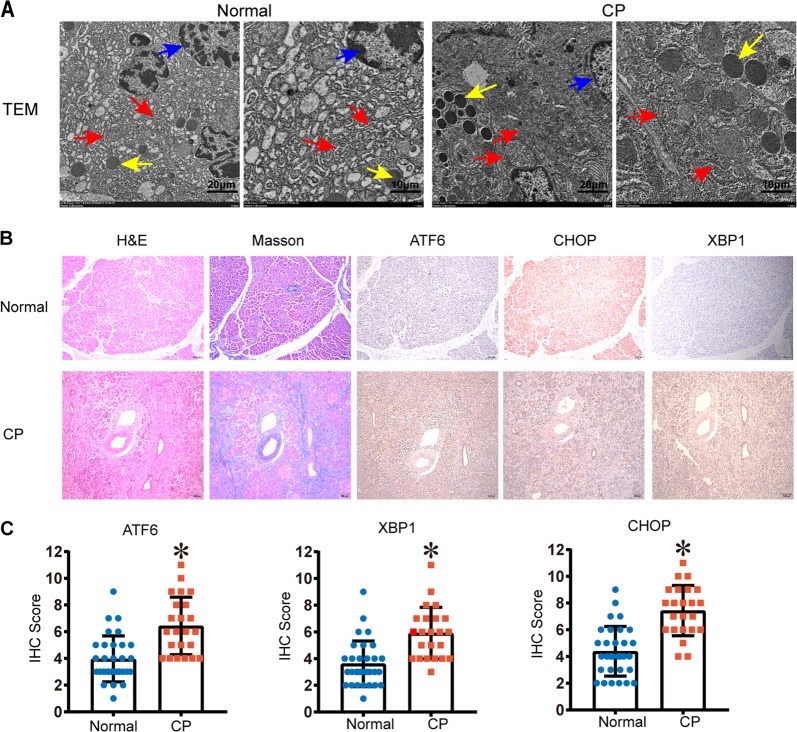
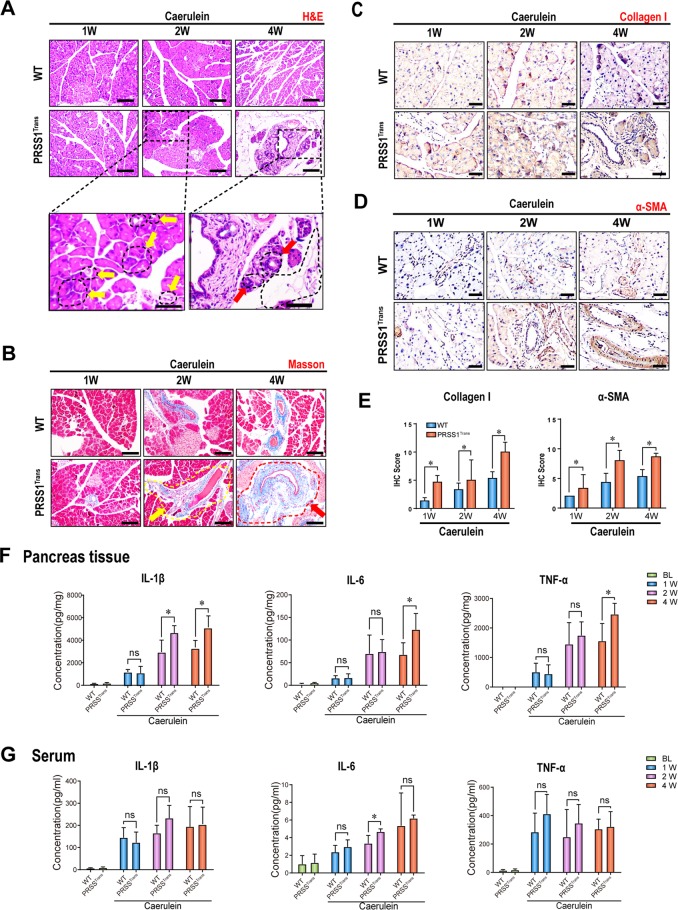
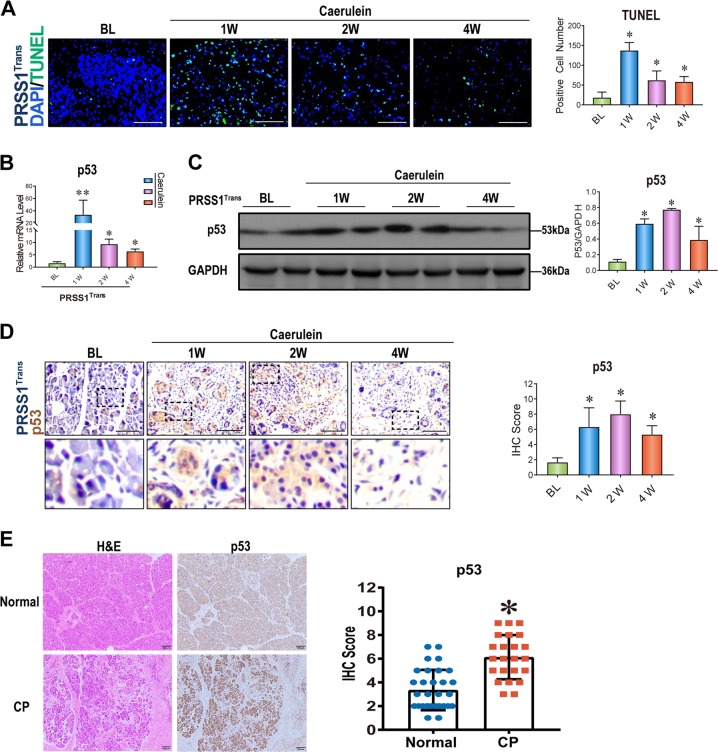
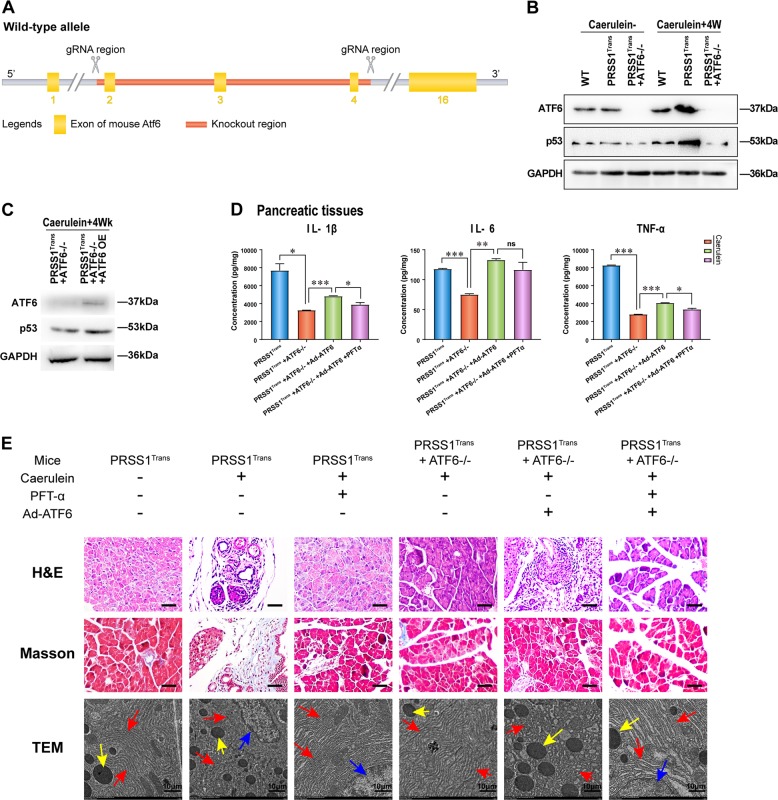
Chronic pancreatitis (CP) is a progressive, recurrent inflammatory disorder of the pancreas. Initiation and progression of CP can result from serine protease 1 (PRSS1) overaccumulation and the ensuing endoplasmic reticulum (ER) stress. However, how ER stress pathways regulate the development and progression of CP remains poorly understood. In the present study we aimed to elucidate the ER stress pathway involved in CP. We found high expression of the ER stress marker genes ATF6, XBP1, and CHOP in human clinical specimens. A humanized PRSS1 transgenic mouse was established and treated with caerulein to mimic the development of CP, as evidenced by pathogenic alterations, collagen deposition, and increased expression of the inflammatory factors IL-6, IL-1β, and TNF-α. ATF6, XBP1, and CHOP expression levels were also increased during CP development in this model. Acinar cell apoptosis was also significantly increased, accompanied by upregulated p53 expression. Inhibition of ATF6 or p53 suppressed the expression of inflammatory factors and progression of CP in the mouse model. Finally, we showed that p53 expression could be regulated by the ATF6/XBP1/CHOP axis to promote the development of CP. We therefore conclude that ATF6 signalling regulates CP progression by modulating pancreatic acinar cell apoptosis, which provides a target for ER stress-based diagnosis and treatment of CP.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



Similar articles
-
ATF6 aggravates acinar cell apoptosis and injury by regulating p53/AIFM2 transcription in Severe Acute Pancreatitis.Theranostics. 2020 Jul 9;10(18):8298-8314. doi: 10.7150/thno.46934. eCollection 2020. Theranostics. 2020. PMID: 32724472 Free PMC article.
-
P53 Activated by ER Stress Aggravates Caerulein-Induced Acute Pancreatitis Progression by Inducing Acinar Cell Apoptosis.Dig Dis Sci. 2020 Nov;65(11):3211-3222. doi: 10.1007/s10620-020-06052-5. Epub 2020 Jan 23. Dig Dis Sci. 2020. PMID: 31974911
-
EMC6 regulates acinar apoptosis via APAF1 in acute and chronic pancreatitis.Cell Death Dis. 2020 Nov 11;11(11):966. doi: 10.1038/s41419-020-03177-3. Cell Death Dis. 2020. PMID: 33177505 Free PMC article.
-
ATF6 upregulates XBP1S and inhibits ER stress-mediated apoptosis in osteoarthritis cartilage.Cell Signal. 2014 Feb;26(2):332-42. doi: 10.1016/j.cellsig.2013.11.018. Epub 2013 Nov 21. Cell Signal. 2014. PMID: 24269637
-
Molecular signal networks and regulating mechanisms of the unfolded protein response.J Zhejiang Univ Sci B. 2017 Jan.;18(1):1-14. doi: 10.1631/jzus.B1600043. J Zhejiang Univ Sci B. 2017. PMID: 28070992 Free PMC article. Review.
Cited by
-
SEC16A Variants Predispose to Chronic Pancreatitis by Impairing ER-to-Golgi Transport and Inducing ER Stress.Adv Sci (Weinh). 2024 Oct;11(38):e2402550. doi: 10.1002/advs.202402550. Epub 2024 Aug 9. Adv Sci (Weinh). 2024. PMID: 39119875 Free PMC article.
-
ATF6α contributes to rheumatoid arthritis by inducing inflammatory cytokine production and apoptosis resistance.Front Immunol. 2022 Oct 10;13:965708. doi: 10.3389/fimmu.2022.965708. eCollection 2022. Front Immunol. 2022. PMID: 36300114 Free PMC article.
-
Immortalized mesenchymal stromal cells overexpressing alpha-1 antitrypsin protect acinar cells from apoptotic and ferroptotic cell death.J Cell Mol Med. 2024 Oct;28(20):e70093. doi: 10.1111/jcmm.70093. J Cell Mol Med. 2024. PMID: 39468387 Free PMC article.
-
Molecular mechanism of ATF6 in unfolded protein response and its role in disease.Heliyon. 2024 Feb 10;10(5):e25937. doi: 10.1016/j.heliyon.2024.e25937. eCollection 2024 Mar 15. Heliyon. 2024. PMID: 38434326 Free PMC article. Review.
-
ATF6 aggravates acinar cell apoptosis and injury by regulating p53/AIFM2 transcription in Severe Acute Pancreatitis.Theranostics. 2020 Jul 9;10(18):8298-8314. doi: 10.7150/thno.46934. eCollection 2020. Theranostics. 2020. PMID: 32724472 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
