

Modeling Human Sexual Motivation in Rodents: Some Caveats
- PMID: 31507386
- PMCID: PMC6719563
- DOI: 10.3389/fnbeh.2019.00187
Modeling Human Sexual Motivation in Rodents: Some Caveats
Abstract
Sexual behavior is activated by motivation. An overwhelming majority of experimental studies of the intricacies of sexual motivation has been performed in rodents, most of them in rats. Sometimes it is desirable to generalize results obtained in this species to other species, particularly the human. It is hoped that studies of the neurobiology of rodent sexual behavior may shed light on the central nervous mechanisms operating in the human, and the search for efficient pharmacological treatments of human sexual dysfunctions relies partly on studies performed in rodents. Then the issue of generalizability of the rodent data to the human becomes crucial. We emphasize the importance of distinguishing between copulatory acts, behavior involving the genitals, and the preceding event, the establishment of physical contact with a potential mate. Comparisons between the structure of copulatory behavior in rats and humans show abysmal differences, but there may be some similarity in the underlying mechanisms. The endocrine control of sex behavior is shortly mentioned, and we also compare the effects of the few drugs known to affect both rodent and human copulatory behavior. The stimuli activating sexual motivation, often called desire in the human literature, are examined, and the sexual approach behaviors in rats and humans are compared. There is a striking similarity between these species in how these behaviors respond to drugs. It is then shown that the intensity of sexual approach is unrelated to the intensity of copulatory behavior. Even though the approach is a requisite for copulation, an activity that requires at least two individuals in close physical contact, these two aspects of sexuality do not covary. This is similar to the role of the testosterone in men and male rats: although the hormone is needed for sex behavior, there is no correlation between serum testosterone concentration and the intensity of copulation. It is also pointed out that human sexual behavior is mostly determined by social conventions, whereas this is not the case in rats and other rodents. It is concluded that some observations in rats can be generalized to the human, but extreme caution must be exercised.
Keywords: ejaculation; lordosis; orgasm; paracopulatory behavior; sexual behavior; sexual motivation.
Figures

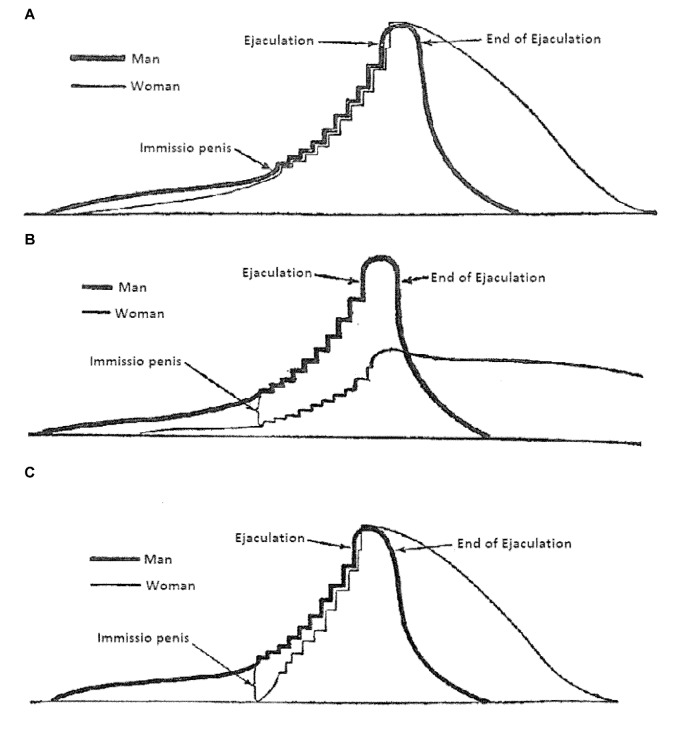
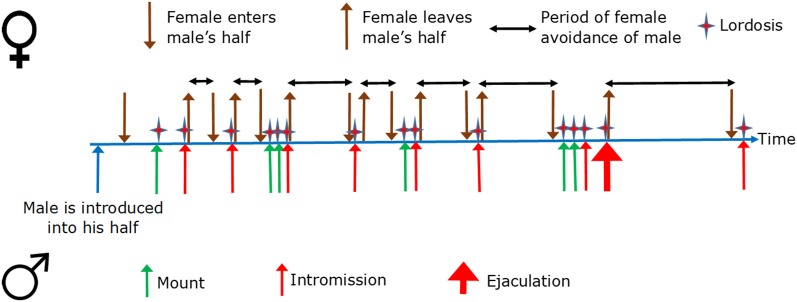
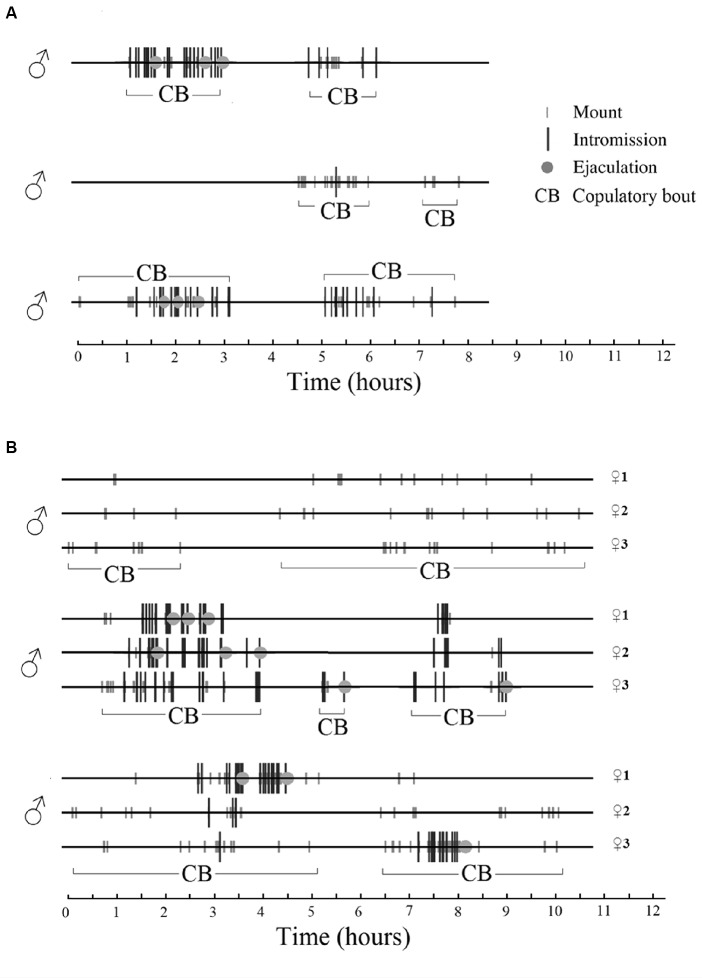
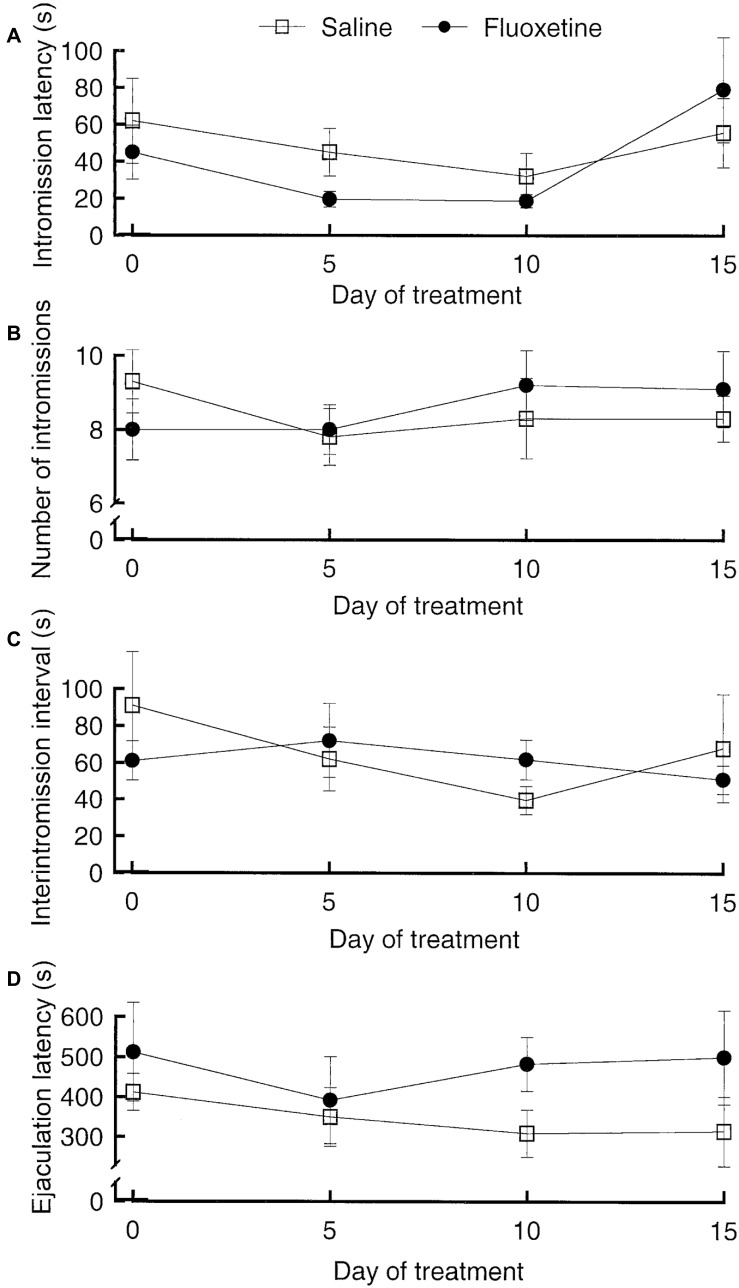
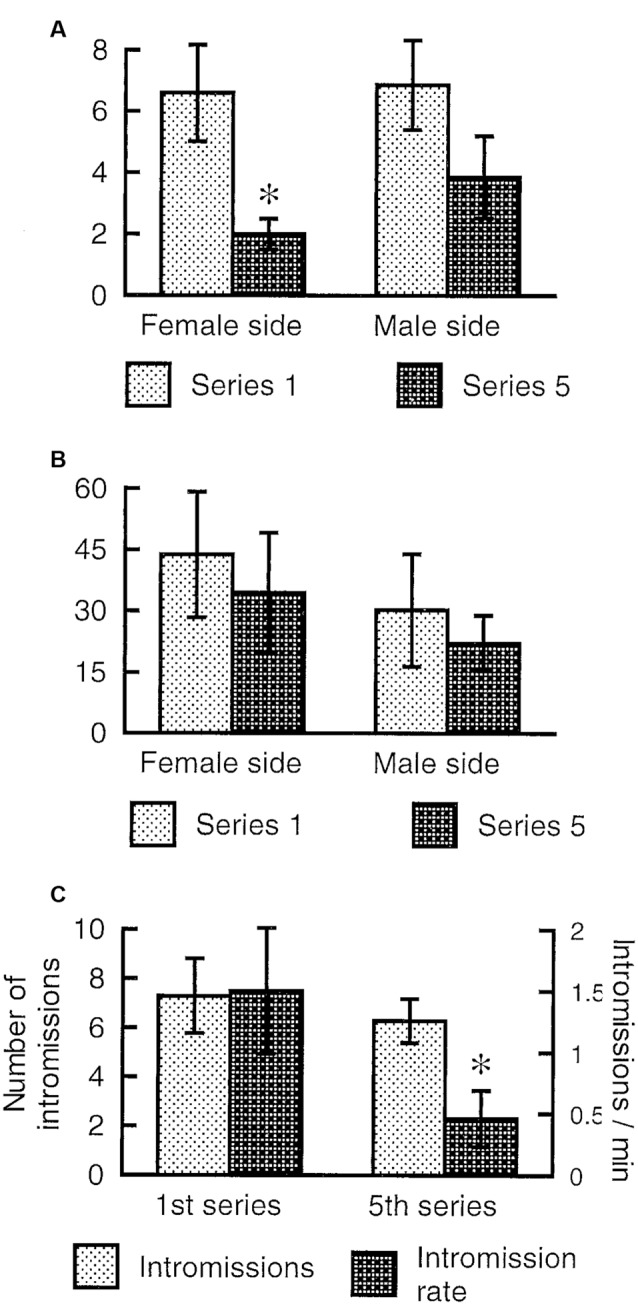

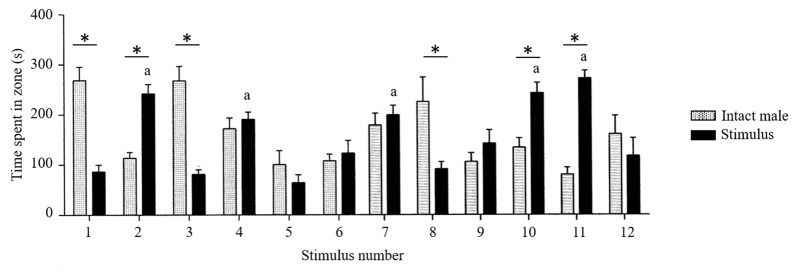
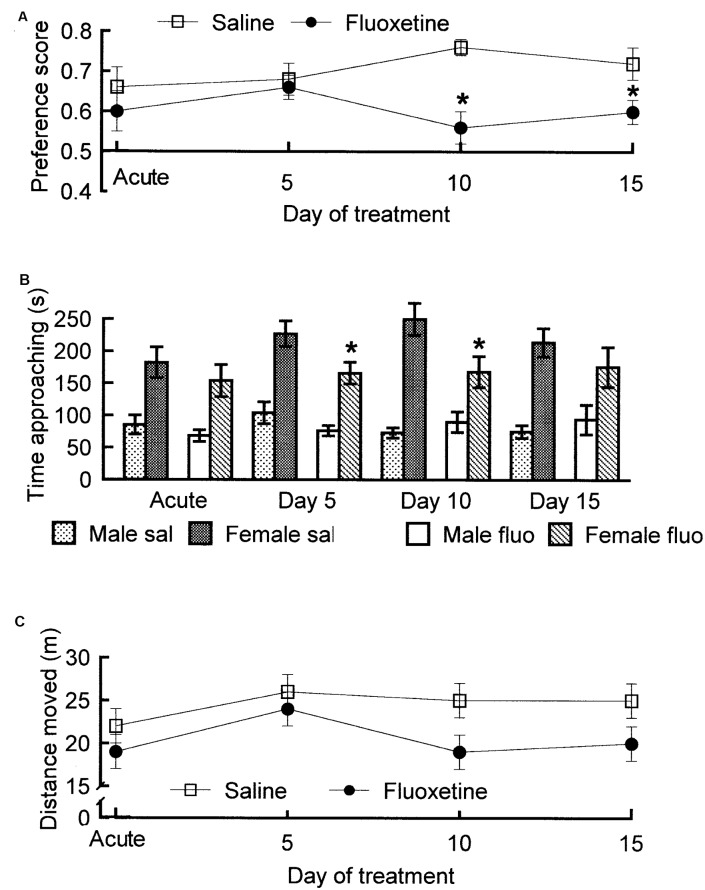
Similar articles
-
The function and meaning of female rat paracopulatory (proceptive) behaviors.Behav Processes. 2015 Sep;118:34-41. doi: 10.1016/j.beproc.2015.05.011. Epub 2015 May 21. Behav Processes. 2015. PMID: 26003137
-
Animal models of female sexual dysfunction: basic considerations on drugs, arousal, motivation and behavior.Pharmacol Biochem Behav. 2014 Jun;121:3-15. doi: 10.1016/j.pbb.2013.10.003. Epub 2013 Oct 11. Pharmacol Biochem Behav. 2014. PMID: 24125786 Review.
-
Effect of dexmedetomidine on ejaculatory behavior and sexual motivation in intact male rats.Pharmacol Biochem Behav. 2012 Dec;103(2):345-52. doi: 10.1016/j.pbb.2012.09.007. Epub 2012 Sep 12. Pharmacol Biochem Behav. 2012. PMID: 22982741
-
Sildenafil and a compound stimulating endothelial NO synthase modify sexual incentive motivation and copulatory behavior in male Wistar and Fisher 344 rats.J Sex Med. 2008 Sep;5(9):2085-99. doi: 10.1111/j.1743-6109.2008.00937.x. Epub 2008 Jul 15. J Sex Med. 2008. PMID: 18637997
-
Sexual incentive motivation, sexual behavior, and general arousal: Do rats and humans tell the same story?Neurosci Biobehav Rev. 2022 Apr;135:104595. doi: 10.1016/j.neubiorev.2022.104595. Epub 2022 Feb 26. Neurosci Biobehav Rev. 2022. PMID: 35231490 Review.
Cited by
-
The Sexual Motivation of Male Rats as a Tool in Animal Models of Human Health Disorders.Front Behav Neurosci. 2019 Nov 26;13:257. doi: 10.3389/fnbeh.2019.00257. eCollection 2019. Front Behav Neurosci. 2019. PMID: 31956302 Free PMC article. Review.
-
Neuroplastic changes in c-Fos, ΔFosB, BDNF, trkB, and Arc expression in the hippocampus of male Roman rats: differential effects of sexual activity.Hippocampus. 2022 Jul;32(7):529-551. doi: 10.1002/hipo.23448. Epub 2022 Jun 18. Hippocampus. 2022. PMID: 35716117 Free PMC article.
-
Sexual Motivation (Desire): Problems with Current Preclinical and Clinical Evaluations of Treatment Effects and a Solution.Behav Sci (Basel). 2025 May 9;15(5):642. doi: 10.3390/bs15050642. Behav Sci (Basel). 2025. PMID: 40426419 Free PMC article. Review.
-
Altered Sexual Behavior in Dopamine Transporter (DAT) Knockout Male Rats: A Behavioral, Neurochemical and Intracerebral Microdialysis Study.Front Behav Neurosci. 2020 Apr 20;14:58. doi: 10.3389/fnbeh.2020.00058. eCollection 2020. Front Behav Neurosci. 2020. PMID: 32372926 Free PMC article.
-
Reduced sensitivity to cocaine effects and changes in mesocorticolimbic dopamine receptors in adolescent sexually active female rats.Psychopharmacology (Berl). 2025 Apr;242(4):817-834. doi: 10.1007/s00213-024-06741-3. Epub 2024 Dec 27. Psychopharmacology (Berl). 2025. PMID: 39729197
References
-
- Ågmo A. (1999). Sexual motivation. An inquiry into events determining the occurrence of sexual behavior. Behav. Brain Res. 105, 129–150. - PubMed
-
- Ågmo A. (2007). Functional and Dysfunctional Sexual Behavior. A Synthesis of Neuroscience and Comparative Psychology. San Diego, CA: Academic Press.
LinkOut - more resources
Full Text Sources