

Specific Sialoforms Required for the Immune Suppressive Activity of Human Soluble CD52
- PMID: 31507595
- PMCID: PMC6719568
- DOI: 10.3389/fimmu.2019.01967
Specific Sialoforms Required for the Immune Suppressive Activity of Human Soluble CD52
Abstract
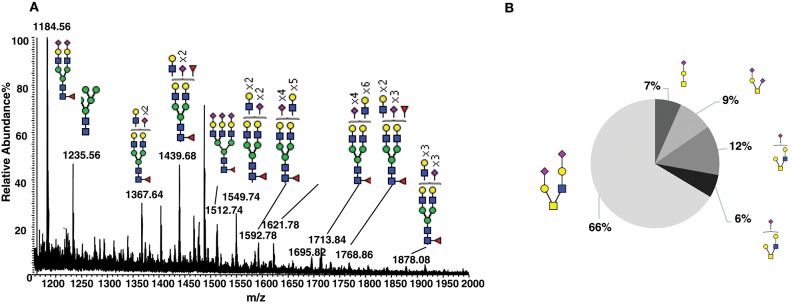
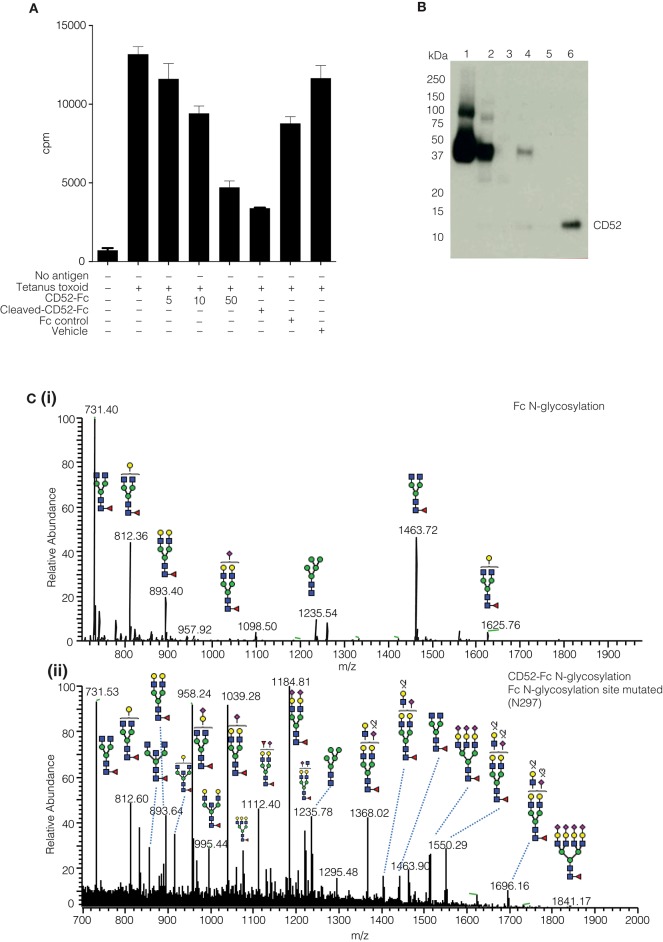
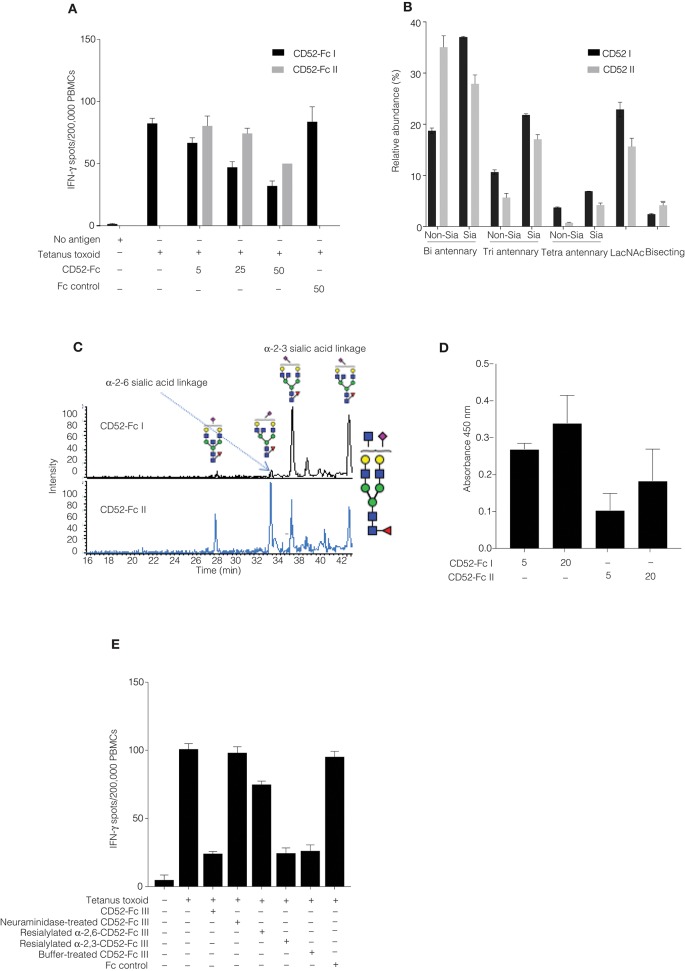
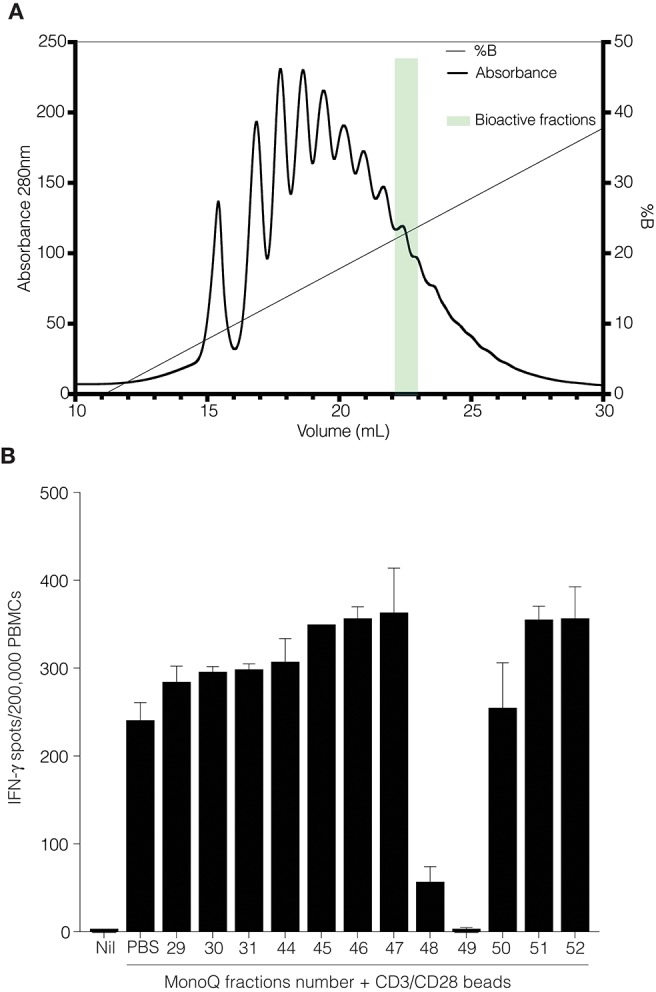
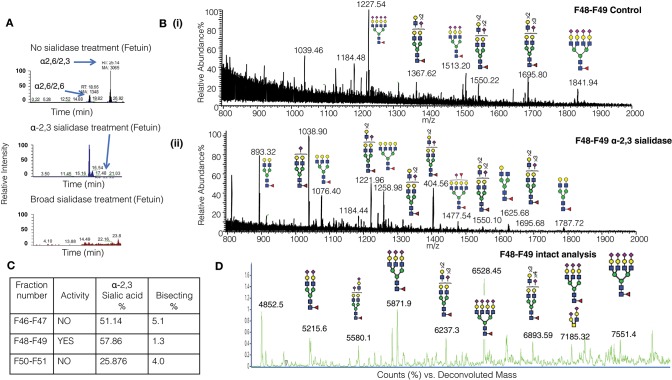
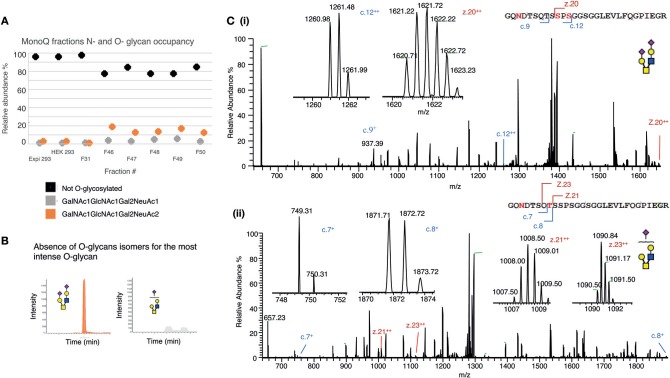
Human CD52 is a small glycopeptide (12 amino acid residues) with one N-linked glycosylation site at asparagine 3 (Asn3) and several potential O-glycosylation serine/threonine sites. Soluble CD52 is released from the surface of activated T cells and mediates immune suppression via its glycan moiety. In suppressing activated T cells, it first sequesters the pro-inflammatory high mobility group Box 1 (HMGB1) protein, which facilitates its binding to the inhibitory sialic acid-binding immunoglobulin-like lectin-10 (Siglec-10) receptor. We aimed to identify the features of CD52 glycan that underlie its bioactivity. Analysis of native CD52 purified from human spleen revealed extensive heterogeneity in N-glycosylation and multi-antennary sialylated N-glycans with abundant polyLacNAc extensions, together with mainly di-sialylated O-glycosylation type structures. Glycomic (porous graphitized carbon-ESI-MS/MS) and glycopeptide (C8-LC-ESI-MS) analysis of recombinant soluble human CD52-immunoglobulin Fc fusion proteins revealed that CD52 bioactivity was correlated with a high abundance of tetra-antennary α-2,3/6 sialylated N-glycans. Removal of α-2,3 sialylation abolished bioactivity, which was restored by re-sialylation with α-2,3 sialyltransferases. When glycoforms of CD52-Fc were fractionated by anion exchange MonoQ-GL chromatography, bioactive fractions displayed mainly tetra-antennary, α-2,3 sialylated N-glycan structures and a lower relative abundance of bisecting GlcNAc structures compared to non-bioactive fractions. In addition, O-glycan core type-2 di-sialylated structures at Ser12 were more abundant in bioactive CD52 fractions. Understanding the structural features of CD52 glycan required for its bioactivity will aid its development as an immunotherapeutic agent.
Keywords: CD52; analysis; glycan structure; immune suppression; tetra-antennary; α-2,3 sialylation.
Figures
Similar articles
-
The molecular basis of immunosuppression by soluble CD52 is defined by interactions of N-linked and O-linked glycans with HMGB1 box B.J Biol Chem. 2025 Apr;301(4):108350. doi: 10.1016/j.jbc.2025.108350. Epub 2025 Feb 25. J Biol Chem. 2025. PMID: 40015632 Free PMC article.
-
CD52 glycan binds the proinflammatory B box of HMGB1 to engage the Siglec-10 receptor and suppress human T cell function.Proc Natl Acad Sci U S A. 2018 Jul 24;115(30):7783-7788. doi: 10.1073/pnas.1722056115. Epub 2018 Jul 11. Proc Natl Acad Sci U S A. 2018. PMID: 29997173 Free PMC article.
-
Understanding of decreased sialylation of Fc-fusion protein in hyperosmotic recombinant Chinese hamster ovary cell culture: N-glycosylation gene expression and N-linked glycan antennary profile.Biotechnol Bioeng. 2017 Aug;114(8):1721-1732. doi: 10.1002/bit.26284. Epub 2017 Mar 30. Biotechnol Bioeng. 2017. PMID: 28266015
-
The immunological function of CD52 and its targeting in organ transplantation.Inflamm Res. 2017 Jul;66(7):571-578. doi: 10.1007/s00011-017-1032-8. Epub 2017 Mar 10. Inflamm Res. 2017. PMID: 28283679 Review.
-
Sialylation of N-glycans: mechanism, cellular compartmentalization and function.Histochem Cell Biol. 2017 Feb;147(2):149-174. doi: 10.1007/s00418-016-1520-x. Epub 2016 Dec 14. Histochem Cell Biol. 2017. PMID: 27975143 Free PMC article. Review.
Cited by
-
Pathogenic T Cells in Celiac Disease Change Phenotype on Gluten Challenge: Implications for T-Cell-Directed Therapies.Adv Sci (Weinh). 2021 Nov;8(21):e2102778. doi: 10.1002/advs.202102778. Epub 2021 Sep 8. Adv Sci (Weinh). 2021. PMID: 34495570 Free PMC article.
-
The Essential Functions and Detection of Bisecting GlcNAc in Cell Biology.Front Chem. 2020 Jul 3;8:511. doi: 10.3389/fchem.2020.00511. eCollection 2020. Front Chem. 2020. PMID: 32719771 Free PMC article. Review.
-
Siglec10-An immunosuppressor and negative predictor of survival prognosis in gliomas.Front Genet. 2022 Nov 16;13:873655. doi: 10.3389/fgene.2022.873655. eCollection 2022. Front Genet. 2022. PMID: 36468012 Free PMC article.
-
Sialoglycan recognition is a common connection linking acidosis, zinc, and HMGB1 in sepsis.Proc Natl Acad Sci U S A. 2021 Mar 9;118(10):e2018090118. doi: 10.1073/pnas.2018090118. Proc Natl Acad Sci U S A. 2021. PMID: 33658363 Free PMC article.
-
Improvement of Lipoplexes With a Sialic Acid Mimetic to Target the C1858T PTPN22 Variant for Immunotherapy in Endocrine Autoimmunity.Front Immunol. 2022 Mar 9;13:838331. doi: 10.3389/fimmu.2022.838331. eCollection 2022. Front Immunol. 2022. PMID: 35355982 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases