

Control of Intestinal Cell Fate by Dynamic Mitotic Spindle Repositioning Influences Epithelial Homeostasis and Longevity
- PMID: 31509744
- PMCID: PMC7046008
- DOI: 10.1016/j.celrep.2019.08.014
Control of Intestinal Cell Fate by Dynamic Mitotic Spindle Repositioning Influences Epithelial Homeostasis and Longevity
Abstract
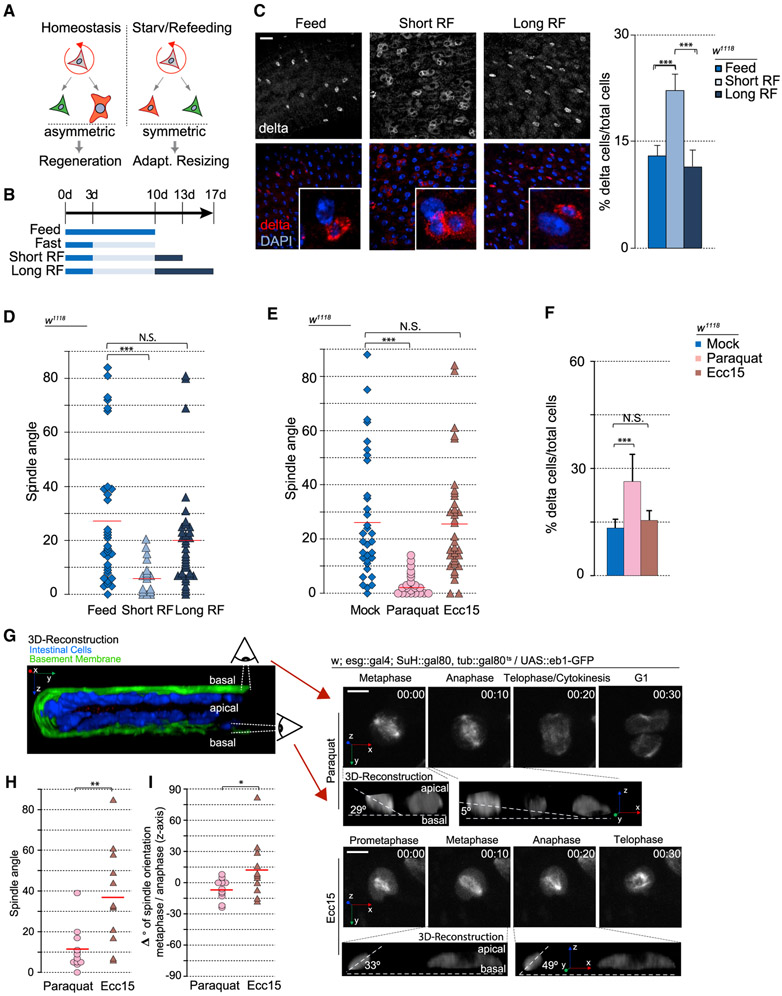
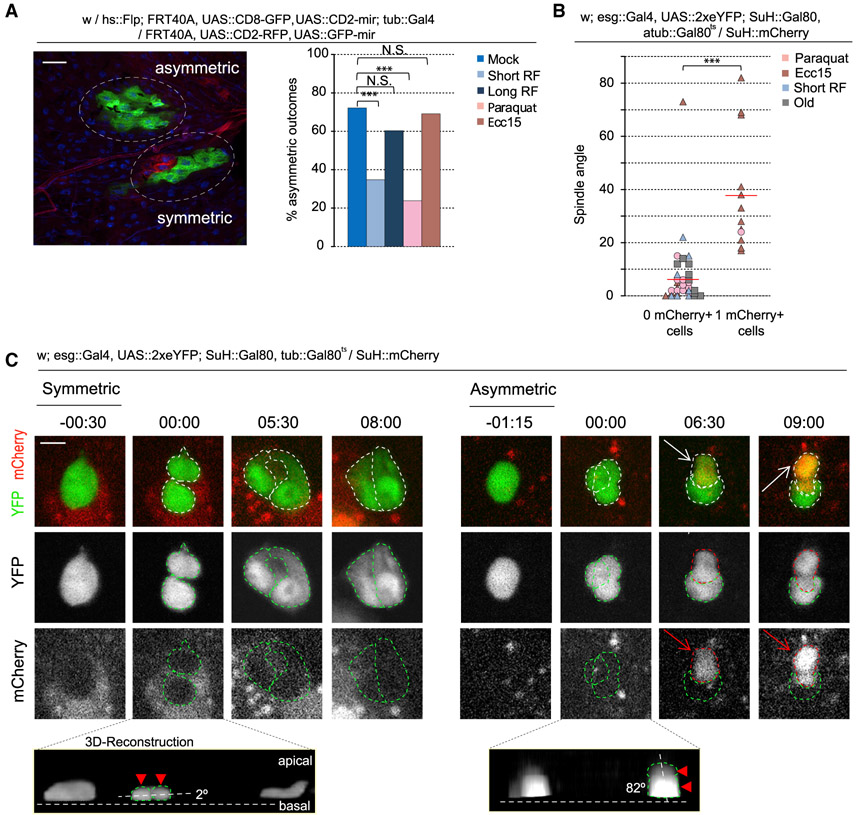
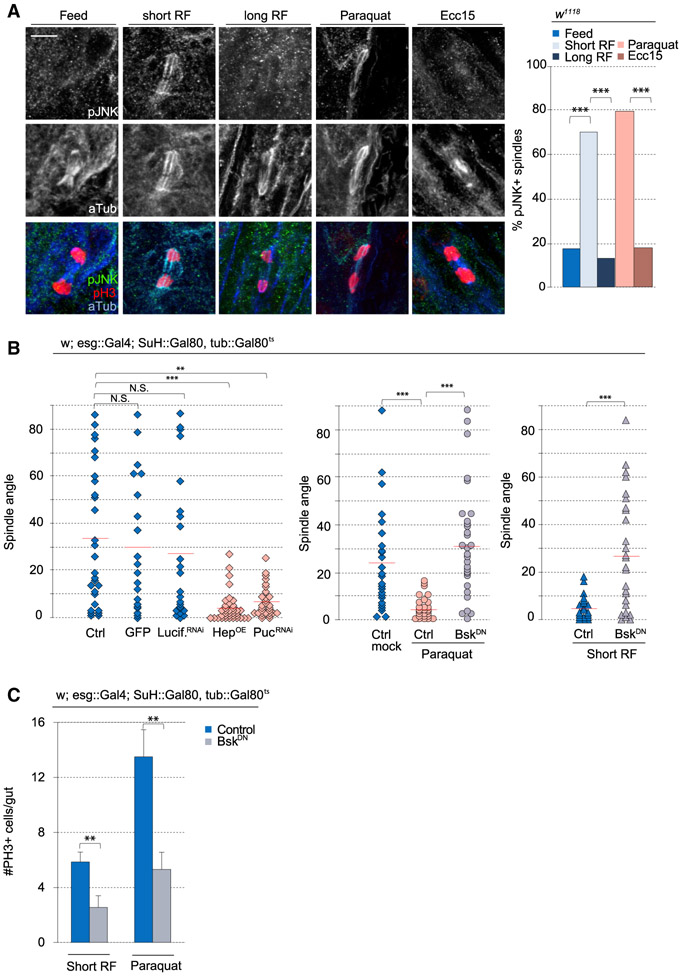
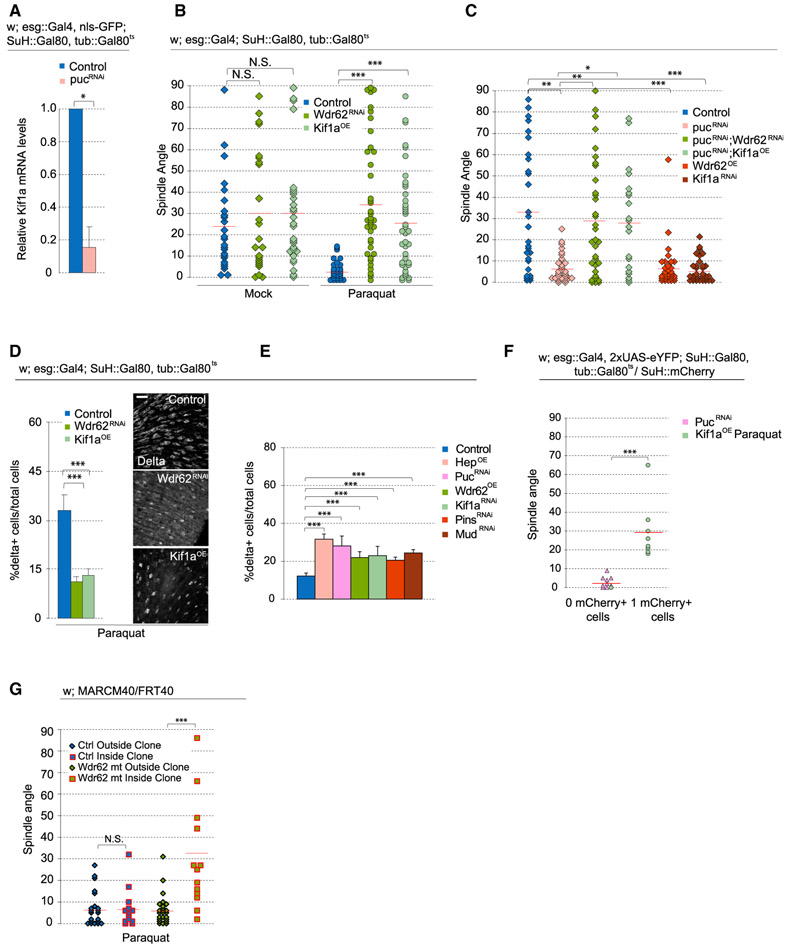
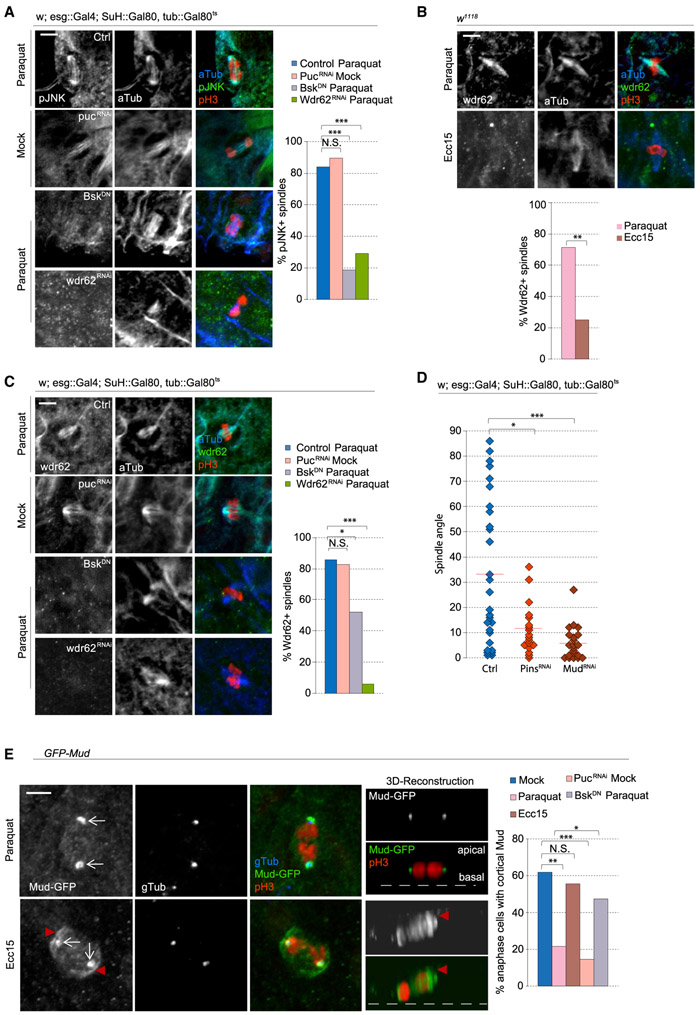
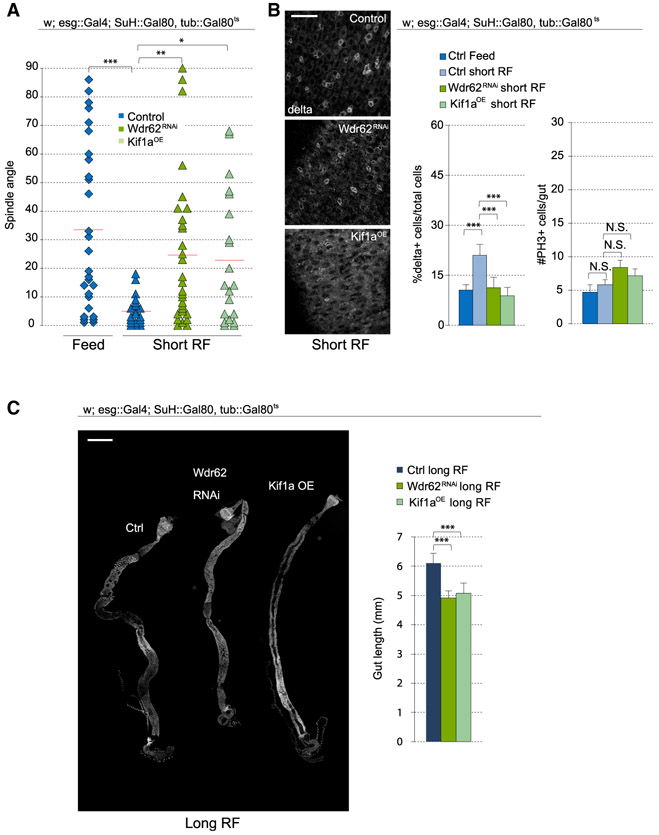
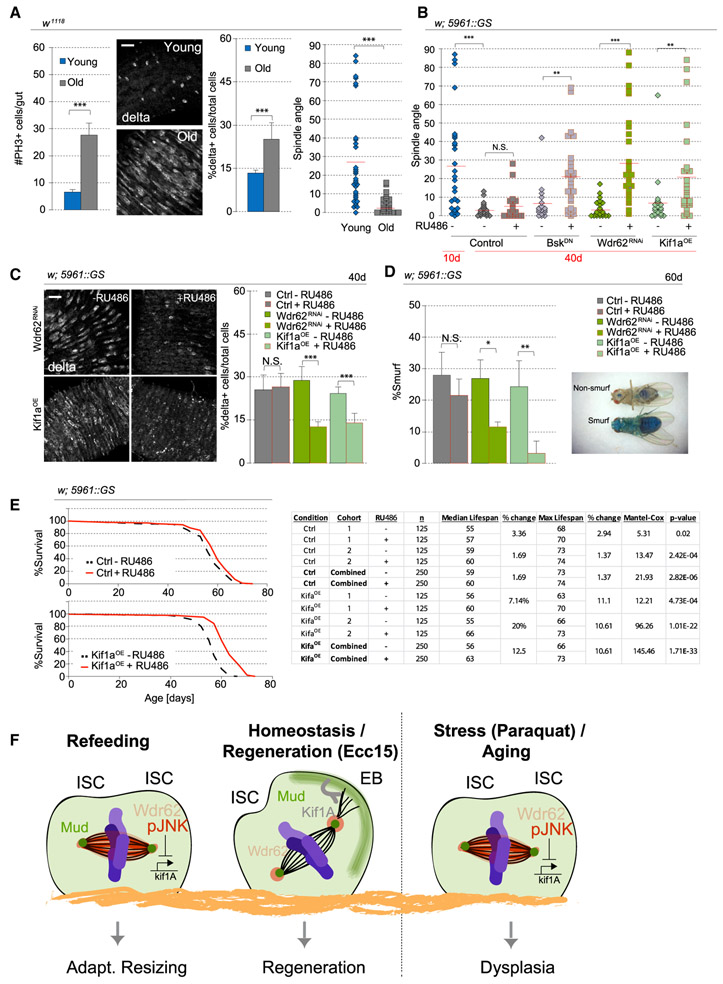
Tissue homeostasis depends on precise yet plastic regulation of stem cell daughter fates. During growth, Drosophila intestinal stem cells (ISCs) adjust fates by switching from asymmetric to symmetric lineages to scale the size of the ISC population. Using a combination of long-term live imaging, lineage tracing, and genetic perturbations, we demonstrate that this switch is executed through the control of mitotic spindle orientation by Jun-N-terminal kinase (JNK) signaling. JNK interacts with the WD40-repeat protein Wdr62 at the spindle and transcriptionally represses the kinesin Kif1a to promote planar spindle orientation. In stress conditions, this function becomes deleterious, resulting in overabundance of symmetric fates and contributing to the loss of tissue homeostasis in the aging animal. Restoring normal ISC spindle orientation by perturbing the JNK/Wdr62/Kif1a axis is sufficient to improve intestinal physiology and extend lifespan. Our findings reveal a critical role for the dynamic control of SC spindle orientation in epithelial maintenance.
Keywords: Drosophila; JNK; Kif1a; Wdr62; aging; cell fate; growth; intestinal stem cell; regeneration; spindle orientation.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Similar articles
-
Lifespan extension by preserving proliferative homeostasis in Drosophila.PLoS Genet. 2010 Oct 14;6(10):e1001159. doi: 10.1371/journal.pgen.1001159. PLoS Genet. 2010. PMID: 20976250 Free PMC article.
-
Drosophila Caliban preserves intestinal homeostasis and lifespan through regulating mitochondrial dynamics and redox state in enterocytes.PLoS Genet. 2020 Oct 15;16(10):e1009140. doi: 10.1371/journal.pgen.1009140. eCollection 2020 Oct. PLoS Genet. 2020. PMID: 33057338 Free PMC article.
-
Asymmetric cell division-dominant neutral drift model for normal intestinal stem cell homeostasis.Am J Physiol Gastrointest Liver Physiol. 2019 Jan 1;316(1):G64-G74. doi: 10.1152/ajpgi.00242.2018. Epub 2018 Oct 25. Am J Physiol Gastrointest Liver Physiol. 2019. PMID: 30359083 Free PMC article.
-
Tissue Adaptation to Environmental Cues by Symmetric and Asymmetric Division Modes of Intestinal Stem Cells.Int J Mol Sci. 2020 Sep 2;21(17):6362. doi: 10.3390/ijms21176362. Int J Mol Sci. 2020. PMID: 32887329 Free PMC article. Review.
-
Promoting longevity by maintaining metabolic and proliferative homeostasis.J Exp Biol. 2014 Jan 1;217(Pt 1):109-18. doi: 10.1242/jeb.089920. J Exp Biol. 2014. PMID: 24353210 Free PMC article. Review.
Cited by
-
Ageing, metabolism and the intestine.EMBO Rep. 2020 Jul 3;21(7):e50047. doi: 10.15252/embr.202050047. Epub 2020 Jun 21. EMBO Rep. 2020. PMID: 32567155 Free PMC article. Review.
-
An improved organ explant culture method reveals stem cell lineage dynamics in the adult Drosophila intestine.Elife. 2022 Aug 25;11:e76010. doi: 10.7554/eLife.76010. Elife. 2022. PMID: 36005292 Free PMC article.
-
Vinculin recruitment to α-catenin halts the differentiation and maturation of enterocyte progenitors to maintain homeostasis of the Drosophila intestine.Elife. 2022 Oct 21;11:e72836. doi: 10.7554/eLife.72836. Elife. 2022. PMID: 36269226 Free PMC article.
-
Drosophila activins adapt gut size to food intake and promote regenerative growth.Nat Commun. 2024 Jan 4;15(1):273. doi: 10.1038/s41467-023-44553-9. Nat Commun. 2024. PMID: 38177201 Free PMC article.
-
The Emerging Roles of JNK Signaling in Drosophila Stem Cell Homeostasis.Int J Mol Sci. 2021 May 24;22(11):5519. doi: 10.3390/ijms22115519. Int J Mol Sci. 2021. PMID: 34073743 Free PMC article. Review.
References
-
- Bellaúche Y, Radovic A, Woods DF, Hough CD, Parmentier ML, O’Kane CJ, Bryant PJ, and Schweisguth F (2001). The Partner of Inscuteable/Discs-large complex is required to establish planar polarity during asymmetric cell division in Drosophila. Cell 106, 355–366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
