

Influenza Viruses in Mice: Deep Sequencing Analysis of Serial Passage and Effects of Sialic Acid Structural Variation
- PMID: 31511393
- PMCID: PMC6854484
- DOI: 10.1128/JVI.01039-19
Influenza Viruses in Mice: Deep Sequencing Analysis of Serial Passage and Effects of Sialic Acid Structural Variation
Abstract
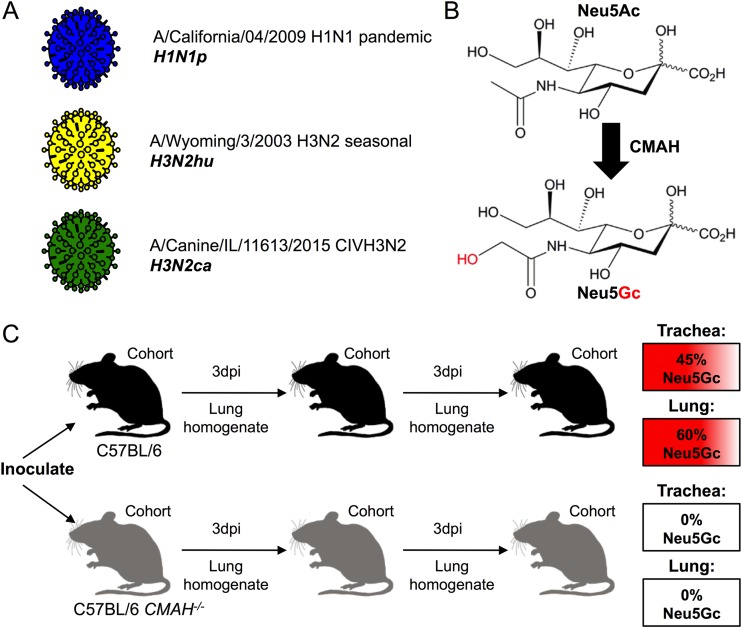
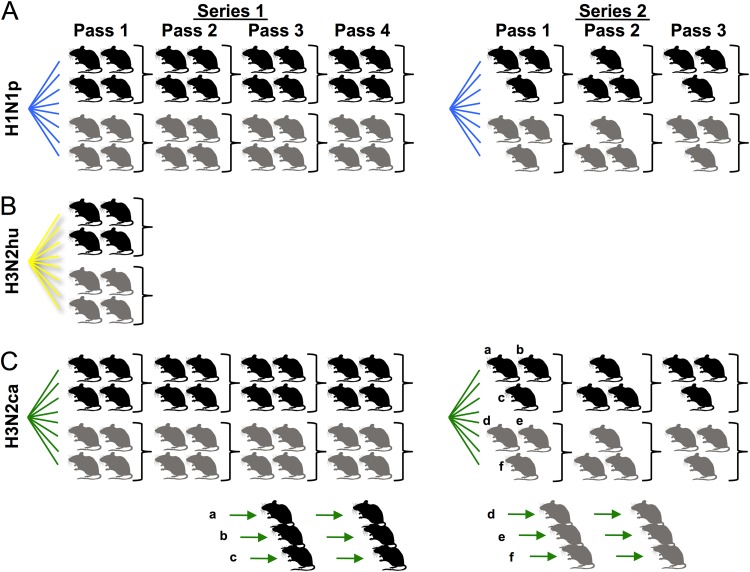
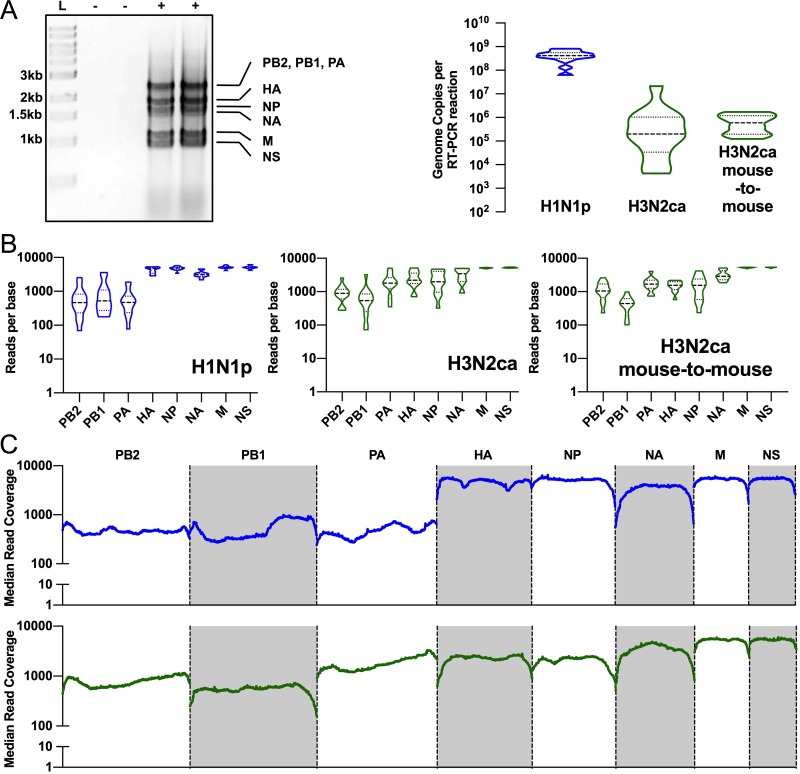
Influenza A viruses have regularly jumped to new host species to cause epidemics or pandemics, an evolutionary process that involves variation in the viral traits necessary to overcome host barriers and facilitate transmission. Mice are not a natural host for influenza virus but are frequently used as models in studies of pathogenesis, often after multiple passages to achieve higher viral titers that result in clinical disease such as weight loss or death. Here, we examine the processes of influenza A virus infection and evolution in mice by comparing single nucleotide variations of a human H1N1 pandemic virus, a seasonal H3N2 virus, and an H3N2 canine influenza virus during experimental passage. We also compared replication and sequence variation in wild-type mice expressing N-glycolylneuraminic acid (Neu5Gc) with those seen in mice expressing only N-acetylneuraminic acid (Neu5Ac). Viruses derived from plasmids were propagated in MDCK cells and then passaged in mice up to four times. Full-genome deep sequencing of the plasmids, cultured viruses, and viruses from mice at various passages revealed only small numbers of mutational changes. The H3N2 canine influenza virus showed increases in frequency of sporadic mutations in the PB2, PA, and NA segments. The H1N1 pandemic virus grew well in mice, and while it exhibited the maintenance of some minority mutations, there was no clear evidence for adaptive evolution. The H3N2 seasonal virus did not establish in the mice. Finally, there were no clear sequence differences associated with the presence or absence of Neu5Gc.IMPORTANCE Mice are commonly used as a model to study the growth and virulence of influenza A viruses in mammals but are not a natural host and have distinct sialic acid receptor profiles compared to humans. Using experimental infections with different subtypes of influenza A virus derived from different hosts, we found that evolution of influenza A virus in mice did not necessarily proceed through the linear accumulation of host-adaptive mutations, that there was variation in the patterns of mutations detected in each repetition, and that the mutation dynamics depended on the virus examined. In addition, variation in the viral receptor, sialic acid, did not affect influenza virus evolution in this model. Overall, our results show that while mice provide a useful animal model for influenza virus pathology, host passage evolution will vary depending on the specific virus tested.
Keywords: animal models; evolution; host range; influenza; sialic acid.
Copyright © 2019 American Society for Microbiology.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
