

Role of laterodorsal tegmentum projections to nucleus accumbens in reward-related behaviors
- PMID: 31515512
- PMCID: PMC6742663
- DOI: 10.1038/s41467-019-11557-3
Role of laterodorsal tegmentum projections to nucleus accumbens in reward-related behaviors
Abstract
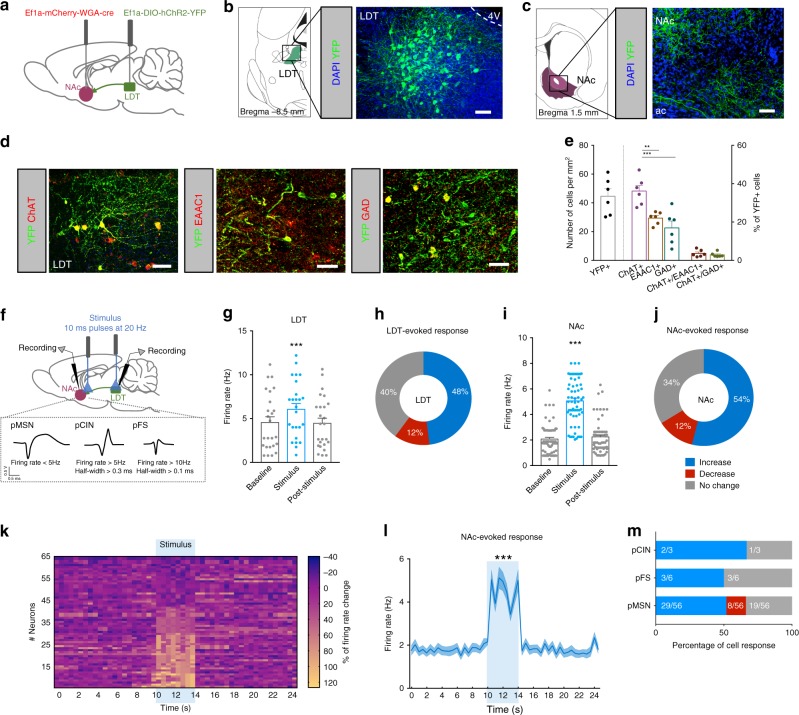
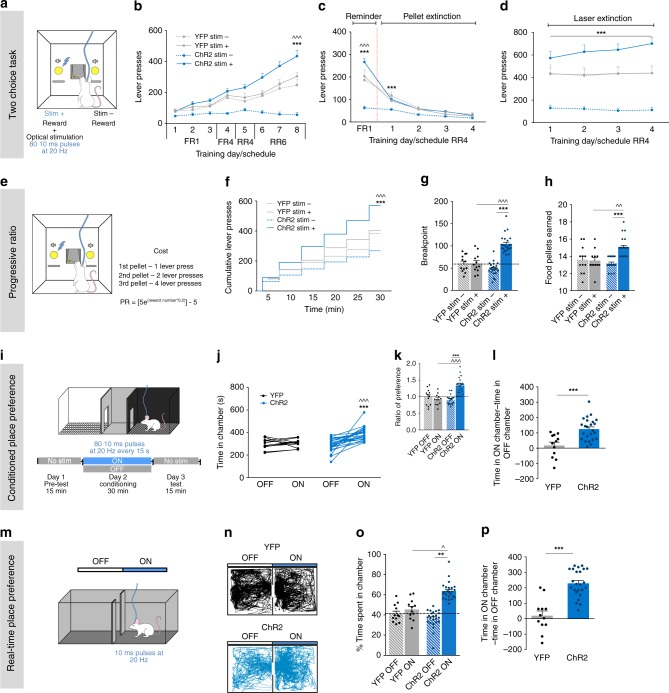
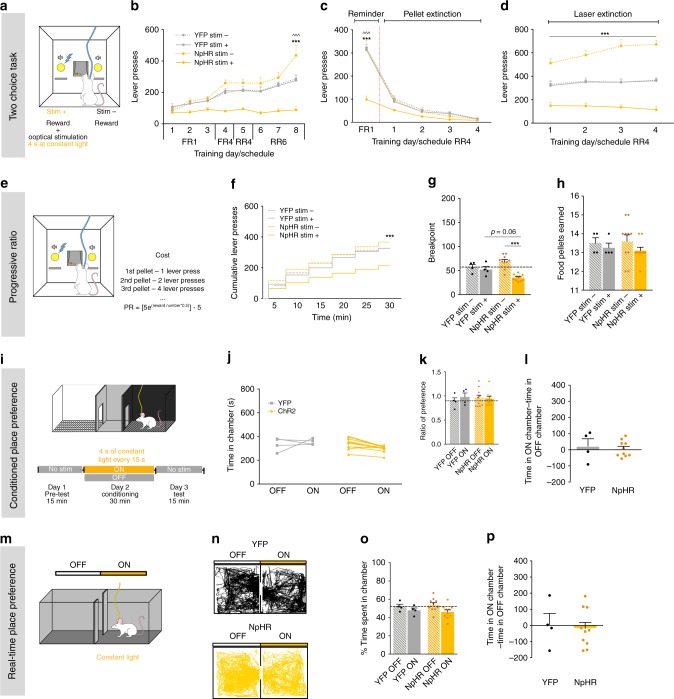
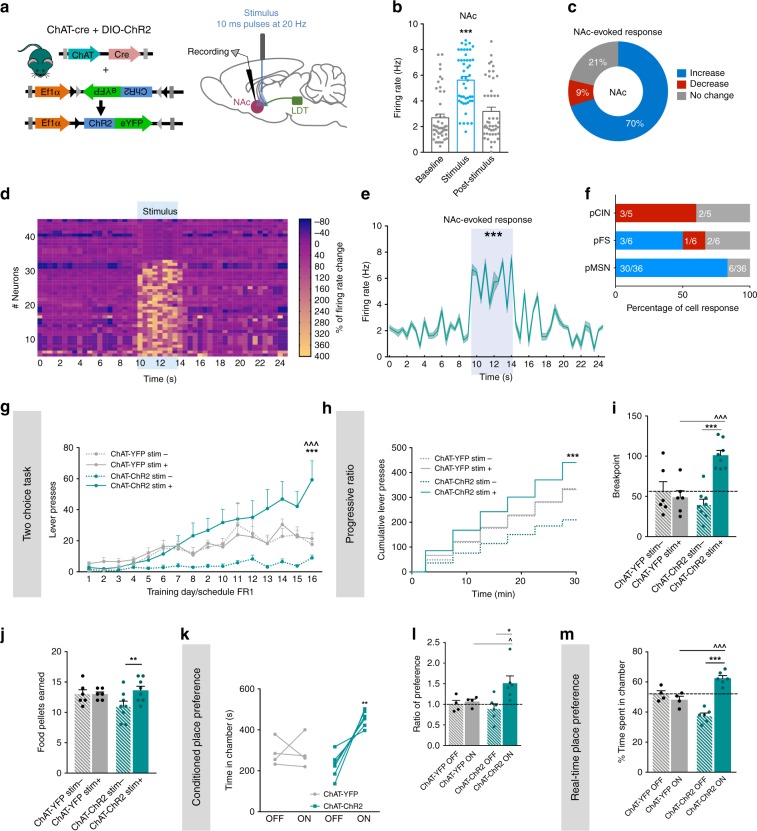
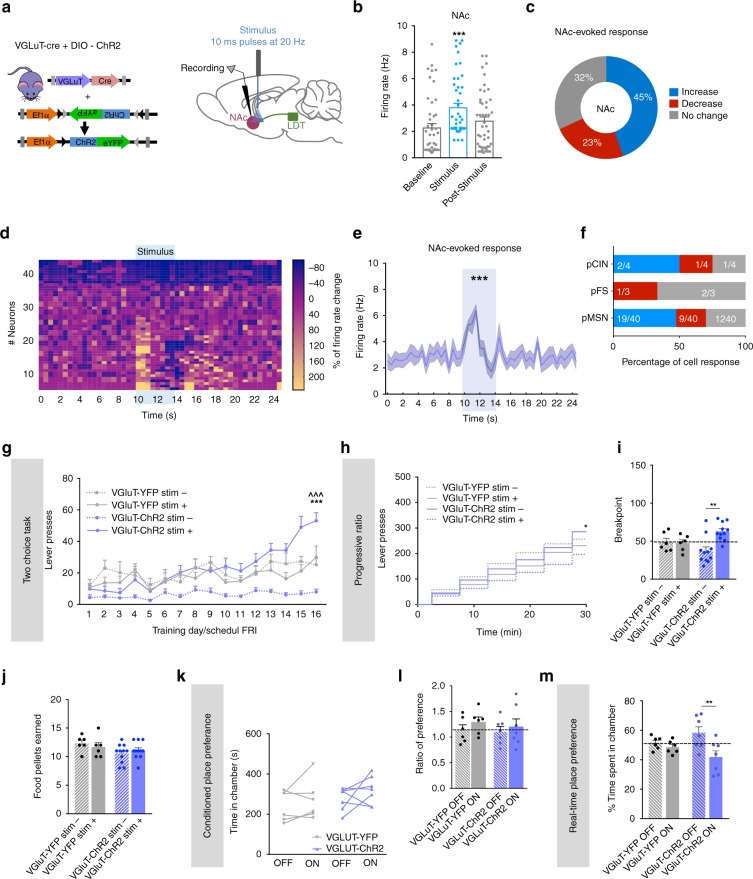
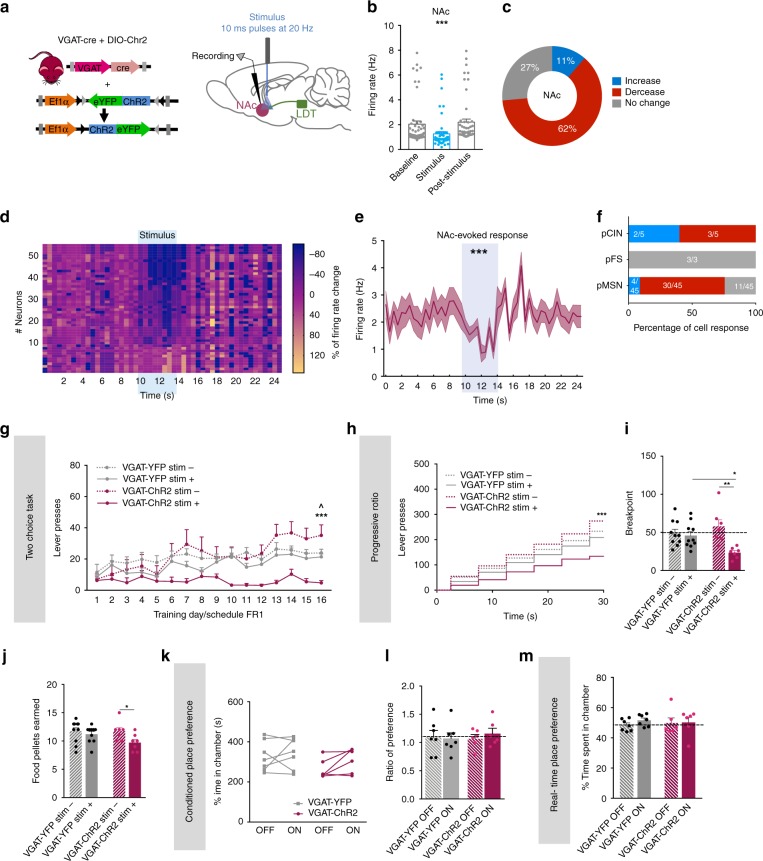
The laterodorsal tegmentum (LDT) is associated with reward considering that it modulates VTA neuronal activity, but recent anatomical evidence shows that the LDT also directly projects to nucleus accumbens (NAc). We show that the majority of LDT-NAc inputs are cholinergic, but there is also GABAergic and glutamatergic innervation; activation of LDT induces a predominantly excitatory response in the NAc. Non-selective optogenetic activation of LDT-NAc projections in rats enhances motivational drive and shifts preference to an otherwise equal reward; whereas inhibition of these projections induces the opposite. Activation of these projections also induces robust place preference. In mice, specific activation of LDT-NAc cholinergic inputs (but not glutamatergic or GABAergic) is sufficient to shift preference, increase motivation, and drive positive reinforcement in different behavioral paradigms. These results provide evidence that LDT-NAc projections play an important role in motivated behaviors and positive reinforcement, and that distinct neuronal populations differentially contribute for these behaviors.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Laterodorsal tegmentum-ventral tegmental area projections encode positive reinforcement signals.J Neurosci Res. 2021 Nov;99(11):3084-3100. doi: 10.1002/jnr.24931. Epub 2021 Aug 10. J Neurosci Res. 2021. PMID: 34374447 Free PMC article.
-
Cholinergic Mesopontine Signals Govern Locomotion and Reward through Dissociable Midbrain Pathways.Neuron. 2016 Apr 20;90(2):333-47. doi: 10.1016/j.neuron.2016.03.028. Neuron. 2016. PMID: 27100197 Free PMC article.
-
Impairments in laterodorsal tegmentum to VTA projections underlie glucocorticoid-triggered reward deficits.Elife. 2017 Aug 24;6:e25843. doi: 10.7554/eLife.25843. Elife. 2017. PMID: 28837419 Free PMC article.
-
Neuroplasticity in cholinergic neurons of the laterodorsal tegmental nucleus contributes to the development of cocaine addiction.Eur J Neurosci. 2019 Aug;50(3):2239-2246. doi: 10.1111/ejn.13962. Epub 2018 Jul 21. Eur J Neurosci. 2019. PMID: 29791036 Review.
-
The role of neuroplasticity in cholinergic neurons of the laterodorsal tegmental nucleus for cocaine addiction.Nihon Arukoru Yakubutsu Igakkai Zasshi. 2016 Oct;51(5):259-67. Nihon Arukoru Yakubutsu Igakkai Zasshi. 2016. PMID: 30478016 Review. English, Japanese.
Cited by
-
Untangling the multifaceted VTA responses to stress.Trends Neurosci. 2025 Aug;48(8):582-593. doi: 10.1016/j.tins.2025.06.004. Epub 2025 Jul 16. Trends Neurosci. 2025. PMID: 40675841 Review.
-
AAV11 enables efficient retrograde targeting of projection neurons and enhances astrocyte-directed transduction.Nat Commun. 2023 Jun 26;14(1):3792. doi: 10.1038/s41467-023-39554-7. Nat Commun. 2023. PMID: 37365155 Free PMC article.
-
Transcription Factors as Important Regulators of Changes in Behavior through Domestication of Gray Rats: Quantitative Data from RNA Sequencing.Int J Mol Sci. 2022 Oct 14;23(20):12269. doi: 10.3390/ijms232012269. Int J Mol Sci. 2022. PMID: 36293128 Free PMC article.
-
A mismatch between striatal cholinergic pauses and dopaminergic reward prediction errors.Proc Natl Acad Sci U S A. 2024 Oct 8;121(41):e2410828121. doi: 10.1073/pnas.2410828121. Epub 2024 Oct 4. Proc Natl Acad Sci U S A. 2024. PMID: 39365823 Free PMC article.
-
Differential cholinergic innervation of lemniscal versus non-lemniscal regions of the inferior colliculus.J Chem Neuroanat. 2024 Sep;139:102443. doi: 10.1016/j.jchemneu.2024.102443. Epub 2024 Jun 22. J Chem Neuroanat. 2024. PMID: 38914378 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
