

Neutrophil Extracellular Traps Drive Mitochondrial Homeostasis in Tumors to Augment Growth
- PMID: 31519688
- PMCID: PMC6825588
- DOI: 10.1158/0008-5472.CAN-19-0800
Neutrophil Extracellular Traps Drive Mitochondrial Homeostasis in Tumors to Augment Growth
Abstract
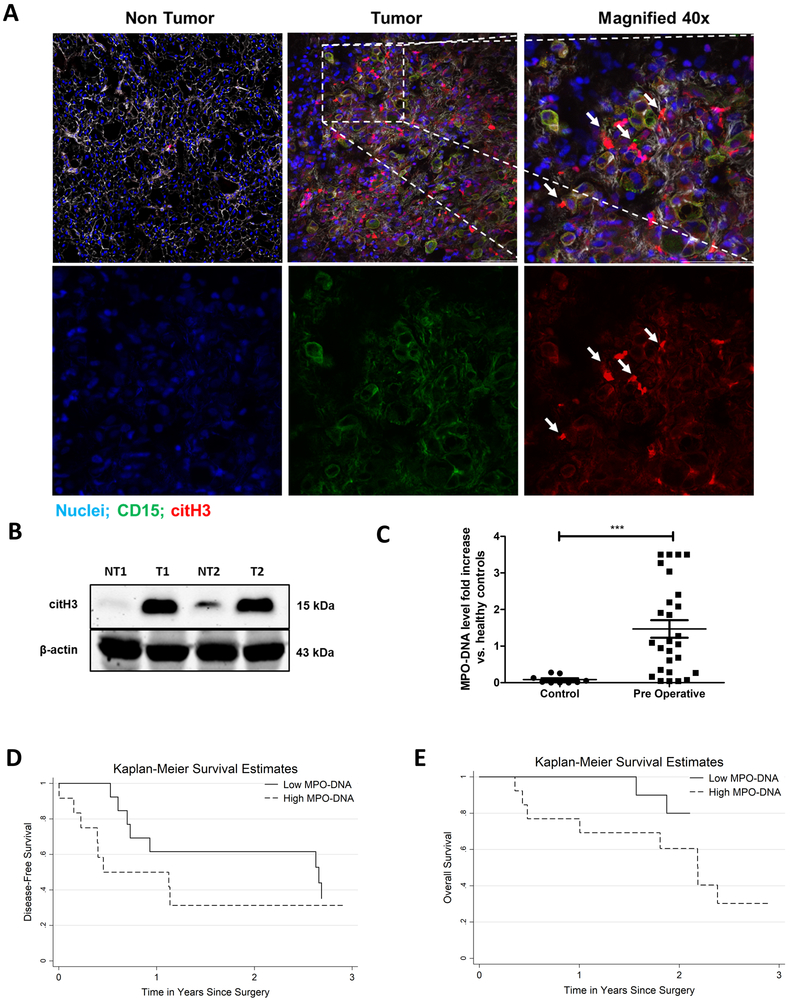
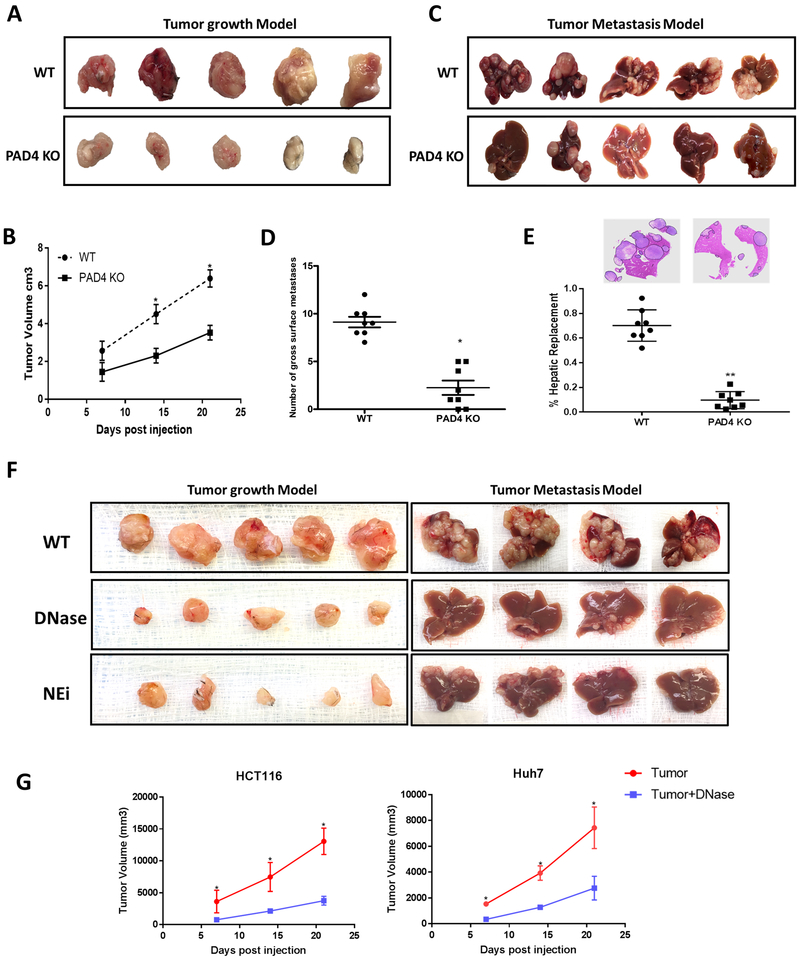
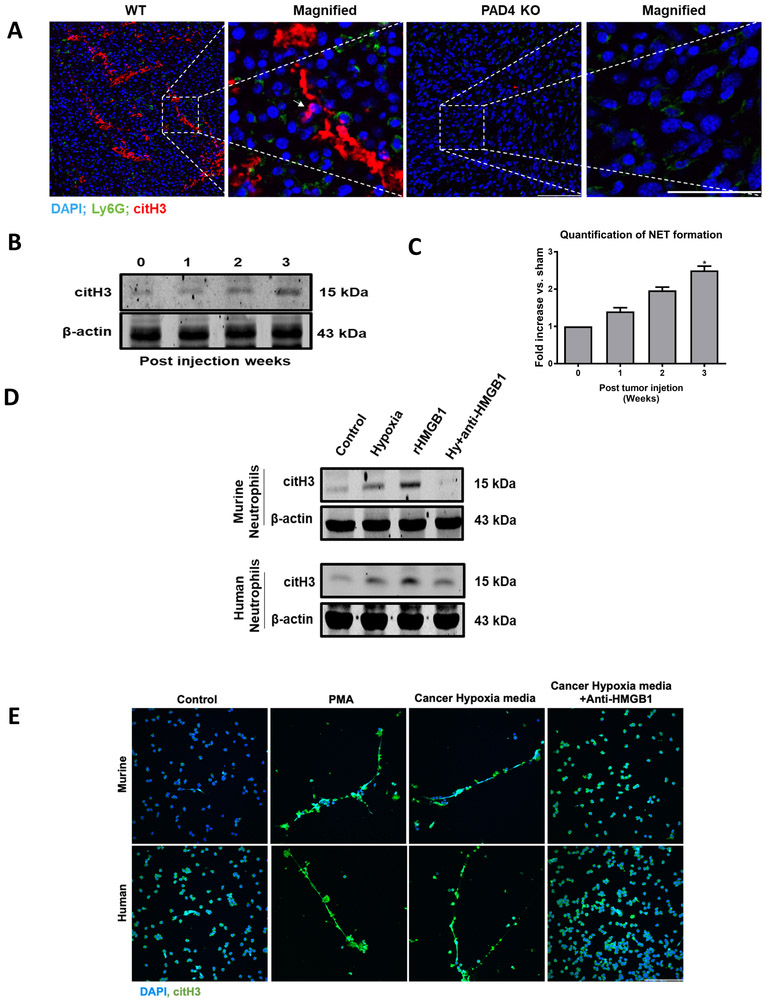
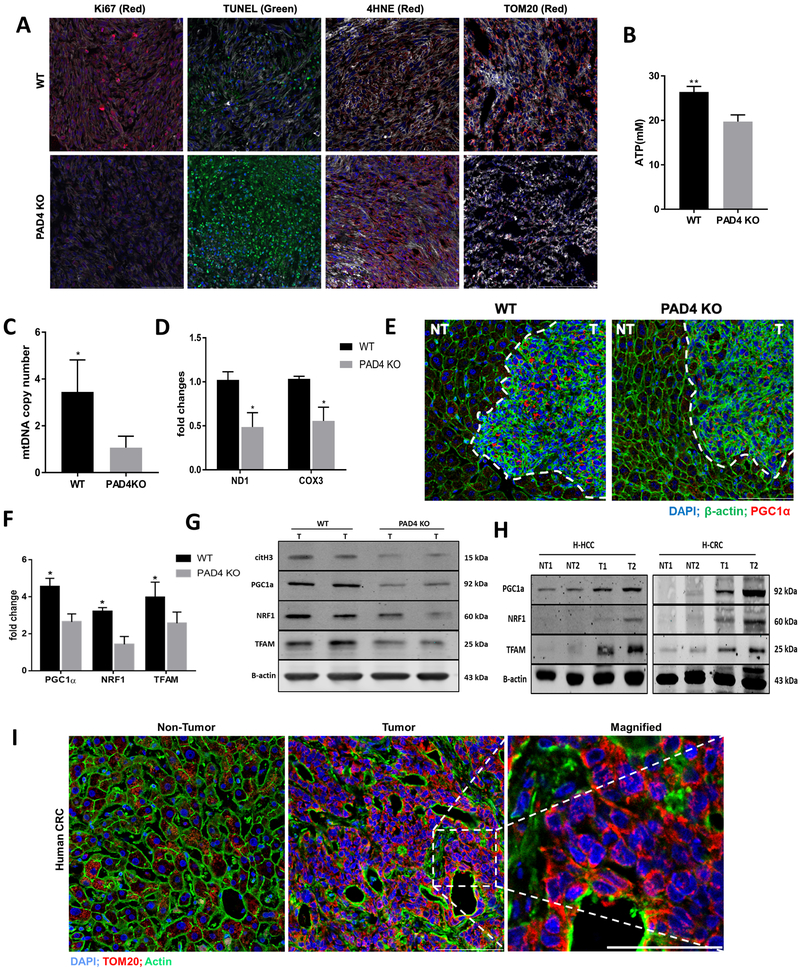
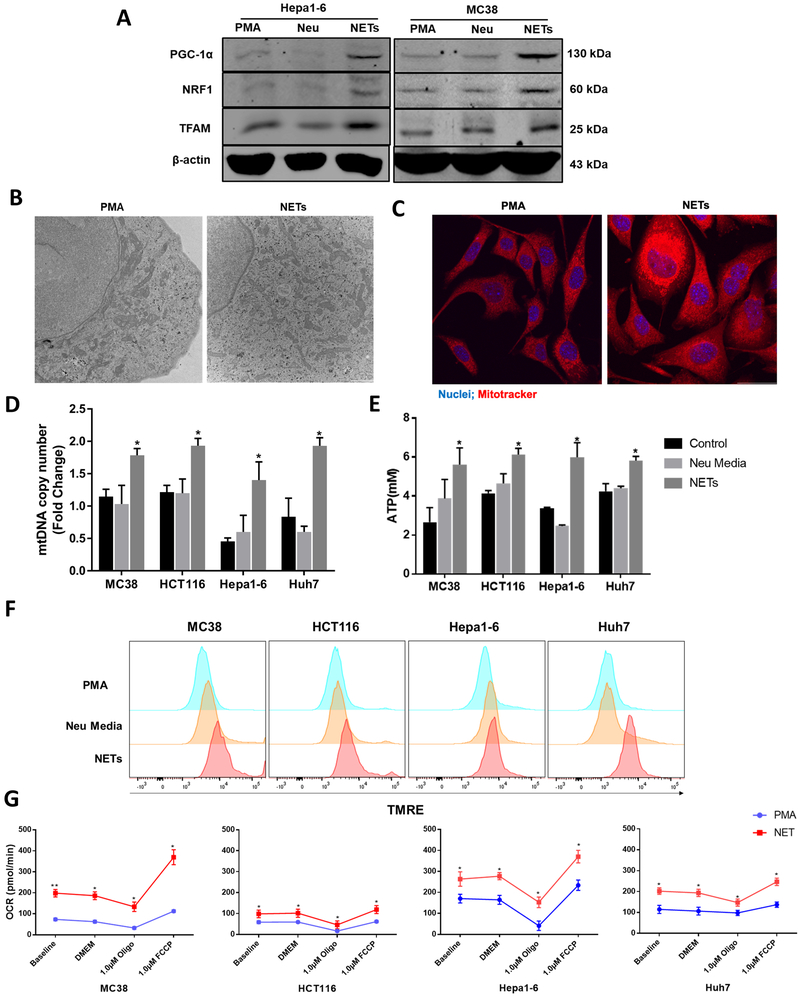
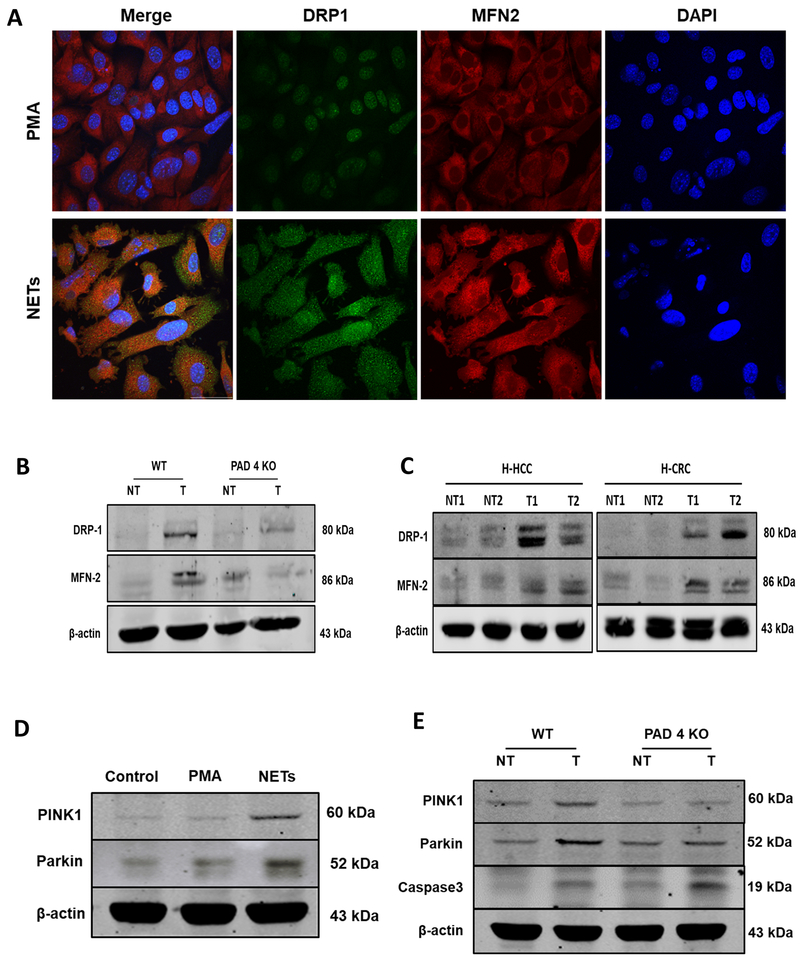
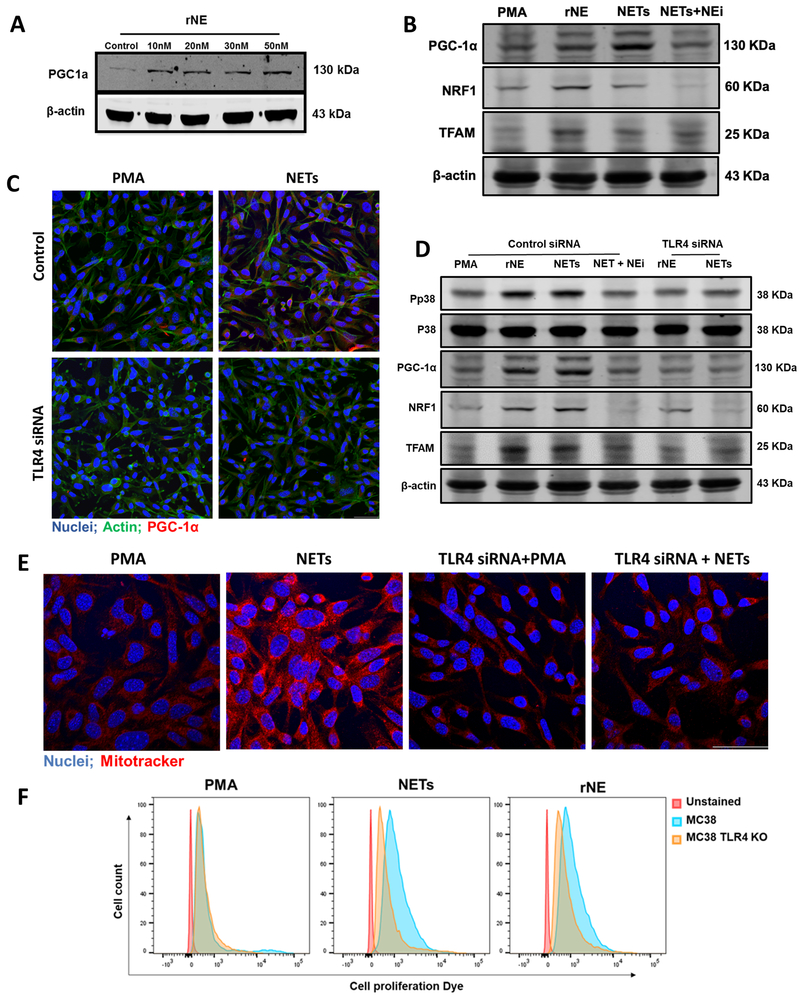
Neutrophil infiltration and neutrophil extracellular traps (NET) in solid cancers are associated with poorer prognosis, but the mechanisms are incompletely understood. We hypothesized that NETs enhance mitochondrial function in tumor cells, providing extra energy for accelerated growth. Metastatic colorectal cancer tissue showed increased intratumoral NETs and supranormal preoperative serum MPO-DNA, a NET marker. Higher MPO-DNA correlated with shorter survival. In mice, subcutaneous tumor implants and hepatic metastases grew slowly in PAD4-KO mice, genetically incapable of NETosis. In parallel experiments, human cancer cell lines grew slower in nu/nu mice treated with DNAse, which disassembles NETs. PAD4-KO tumors manifested decreased proliferation, increased apoptosis, and increased evidence of oxidative stress. PAD4-KO tumors had decreased mitochondrial density, mitochondrial DNA, a lesser degree of ATP production, along with significantly decreased mitochondrial biogenesis proteins PGC1α, TFAM, and NRF-1. In vitro, cancer cells treated with NETs upregulated mitochondrial biogenesis-associated genes, increased mitochondrial density, increased ATP production, enhanced the percentage of cancer cells with reduced mitochondrial membrane potential, and increased the oxygen consumption rate. Furthermore, NETs increased cancer cells' expression of fission and fusion-associated proteins, DRP-1 and MFN-2, and mitophagy-linked proteins, PINK1 and Parkin. All of which were decreased in PAD4-KO tumors. Mechanistically, neutrophil elastase released from NETs activated TLR4 on cancer cells, leading to PGC1α upregulation, increased mitochondrial biogenesis, and accelerated growth. Taken together, NETs can directly alter the metabolic programming of cancer cells to increase tumor growth. NETs represent a promising therapeutic target to halt cancer progression. SIGNIFICANCE: Neutrophils through the release of NETs facilitate the growth of stressed cancer cells by altering their bioenergetics, the inhibition of which induces cell death.
©2019 American Association for Cancer Research.
Conflict of interest statement
Figures
References
-
- Li YW, Qiu SJ, Fan J, Zhou J, Gao Q, Xiao YS, et al. Intratumoral neutrophils: A poor prognostic factor for hepatocellular carcinoma following resection. J Hepatol. 2011; - PubMed
-
- J TO, S H, M HJ, D F, H M, S P, et al. Intratumoral neutrophils, plasmacytoid dendritic cells, and pSTAT3 in AJCC stage I/II melanoma prognosis. J. Clin. Oncol. 2011. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
