

DNA Sequence Is a Major Determinant of Tetrasome Dynamics
- PMID: 31521330
- PMCID: PMC6895708
- DOI: 10.1016/j.bpj.2019.07.055
DNA Sequence Is a Major Determinant of Tetrasome Dynamics
Abstract
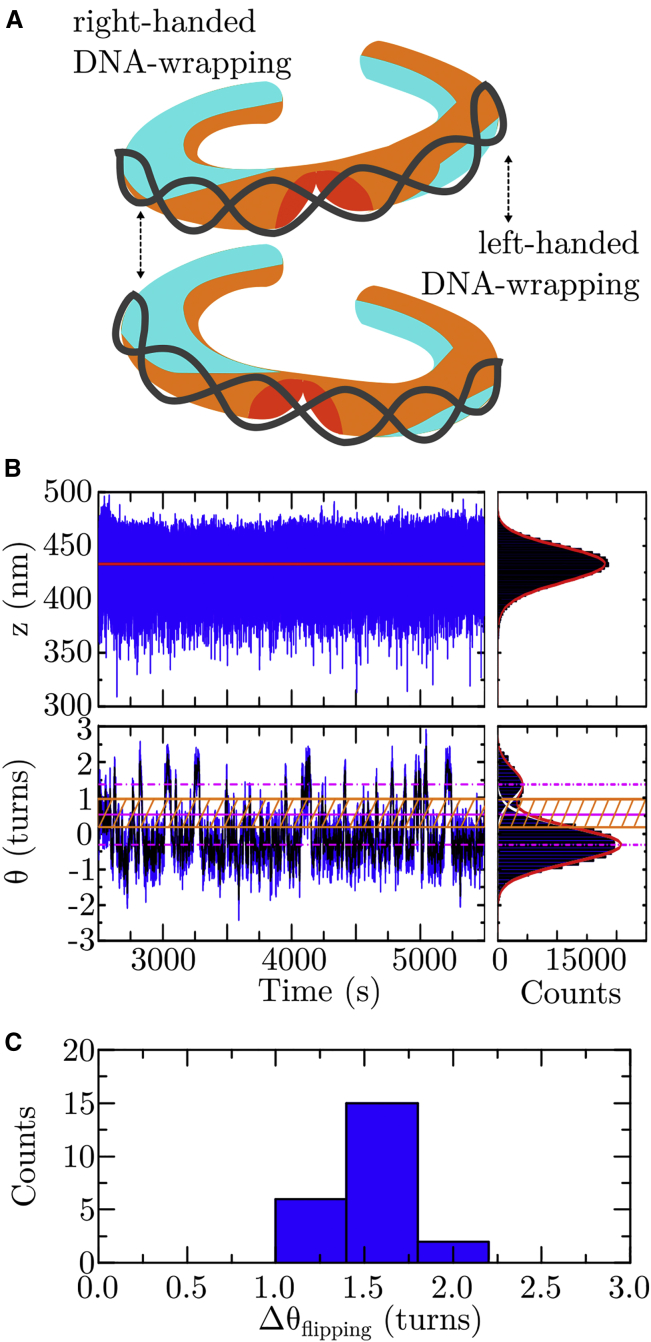
Eukaryotic genomes are hierarchically organized into protein-DNA assemblies for compaction into the nucleus. Nucleosomes, with the (H3-H4)2 tetrasome as a likely intermediate, are highly dynamic in nature by way of several different mechanisms. We have recently shown that tetrasomes spontaneously change the direction of their DNA wrapping between left- and right-handed conformations, which may prevent torque buildup in chromatin during active transcription or replication. DNA sequence has been shown to strongly affect nucleosome positioning throughout chromatin. It is not known, however, whether DNA sequence also impacts the dynamic properties of tetrasomes. To address this question, we examined tetrasomes assembled on a high-affinity DNA sequence using freely orbiting magnetic tweezers. In this context, we also studied the effects of mono- and divalent salts on the flipping dynamics. We found that neither DNA sequence nor altered buffer conditions affect overall tetrasome structure. In contrast, tetrasomes bound to high-affinity DNA sequences showed significantly altered flipping kinetics, predominantly via a reduction in the lifetime of the canonical state of left-handed wrapping. Increased mono- and divalent salt concentrations counteracted this behavior. Thus, our study indicates that high-affinity DNA sequences impact not only the positioning of the nucleosome but that they also endow the subnucleosomal tetrasome with enhanced conformational plasticity. This may provide a means to prevent histone loss upon exposure to torsional stress, thereby contributing to the integrity of chromatin at high-affinity sites.
Copyright © 2019 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures


Similar articles
-
Comparing the Assembly and Handedness Dynamics of (H3.3-H4)2 Tetrasomes to Canonical Tetrasomes.PLoS One. 2015 Oct 27;10(10):e0141267. doi: 10.1371/journal.pone.0141267. eCollection 2015. PLoS One. 2015. PMID: 26506534 Free PMC article.
-
Modification of the histone tetramer at the H3-H3 interface impacts tetrasome conformations and dynamics.J Chem Phys. 2018 Mar 28;148(12):123323. doi: 10.1063/1.5009100. J Chem Phys. 2018. PMID: 29604863
-
Nucleosome dynamics. Protein and DNA contributions in the chiral transition of the tetrasome, the histone (H3-H4)2 tetramer-DNA particle.J Mol Biol. 1999 Aug 27;291(4):815-41. doi: 10.1006/jmbi.1999.2988. J Mol Biol. 1999. PMID: 10452891
-
Nucleosome conformational flexibility and implications for chromatin dynamics.Philos Trans A Math Phys Eng Sci. 2004 Jul 15;362(1820):1519-47. doi: 10.1098/rsta.2004.1387. Philos Trans A Math Phys Eng Sci. 2004. PMID: 15306464 Review.
-
DNA recognition and nucleosome organization.Biopolymers. 1997;44(4):423-33. doi: 10.1002/(SICI)1097-0282(1997)44:4<423::AID-BIP6>3.0.CO;2-M. Biopolymers. 1997. PMID: 9782778 Review.
Cited by
-
DNAcycP: a deep learning tool for DNA cyclizability prediction.Nucleic Acids Res. 2022 Apr 8;50(6):3142-3154. doi: 10.1093/nar/gkac162. Nucleic Acids Res. 2022. PMID: 35288750 Free PMC article.
-
The esBAF and ISWI nucleosome remodeling complexes influence occupancy of overlapping dinucleosomes and fragile nucleosomes in murine embryonic stem cells.BMC Genomics. 2023 Apr 13;24(1):201. doi: 10.1186/s12864-023-09287-4. BMC Genomics. 2023. PMID: 37055726 Free PMC article.
-
Histone chaperone-mediated co-expression assembly of tetrasomes and nucleosomes.FEBS Open Bio. 2021 Nov;11(11):2912-2920. doi: 10.1002/2211-5463.13311. Epub 2021 Oct 19. FEBS Open Bio. 2021. PMID: 34614293 Free PMC article.
-
Histone tetrasome dynamics affects chromatin transcription.Nucleic Acids Res. 2025 Apr 22;53(8):gkaf356. doi: 10.1093/nar/gkaf356. Nucleic Acids Res. 2025. PMID: 40304183 Free PMC article.
-
Brewing COFFEE: A sequence-specific coarse-grained energy function for simulations of DNA-protein complexes.bioRxiv [Preprint]. 2023 Aug 1:2023.06.07.544064. doi: 10.1101/2023.06.07.544064. bioRxiv. 2023. Update in: J Chem Theory Comput. 2024 Feb 13;20(3):1398-1413. doi: 10.1021/acs.jctc.3c00833. PMID: 37333386 Free PMC article. Updated. Preprint.
References
-
- Kornberg R.D. Chromatin structure: a repeating unit of histones and DNA. Science. 1974;184:868–871. - PubMed
-
- Olins A.L., Olins D.E. Spheroid chromatin units (v bodies) Science. 1974;183:330–332. - PubMed
-
- Oudet P., Gross-Bellard M., Chambon P. Electron microscopic and biochemical evidence that chromatin structure is a repeating unit. Cell. 1975;4:281–300. - PubMed
-
- Richmond T.J., Finch J.T., Klug A. Structure of the nucleosome core particle at 7 A resolution. Nature. 1984;311:532–537. - PubMed
-
- Luger K., Mäder A.W., Richmond T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
