

Robust, Long-Term Culture of Endoderm-Derived Hepatic Organoids for Disease Modeling
- PMID: 31522975
- PMCID: PMC6829764
- DOI: 10.1016/j.stemcr.2019.08.007
Robust, Long-Term Culture of Endoderm-Derived Hepatic Organoids for Disease Modeling
Abstract
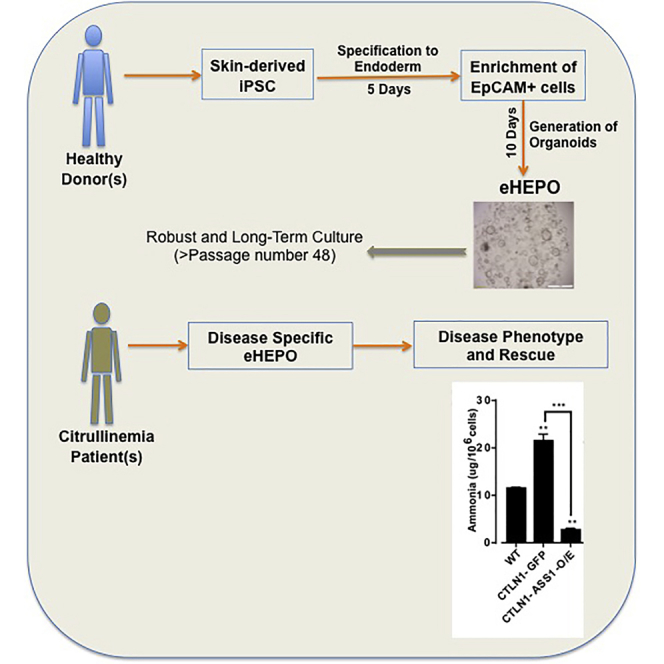
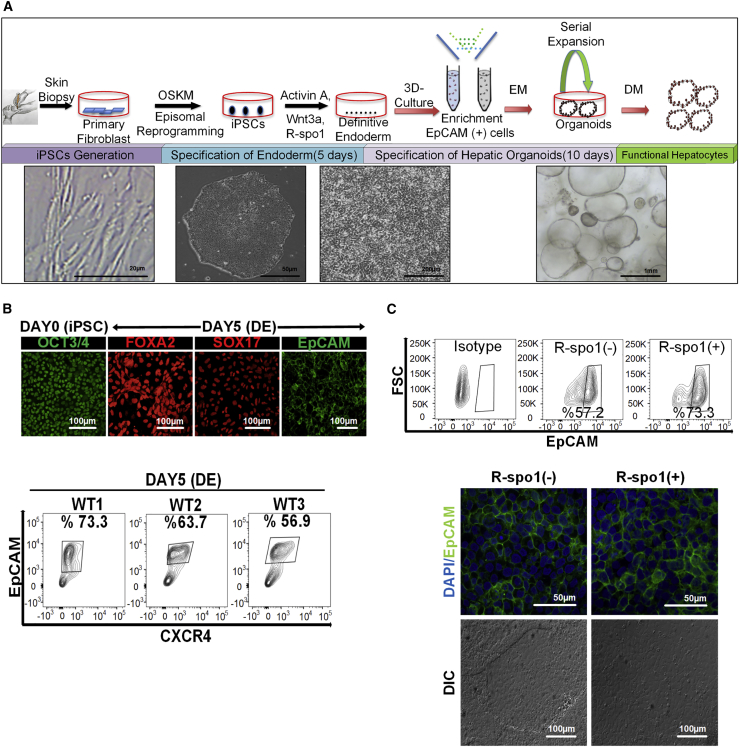
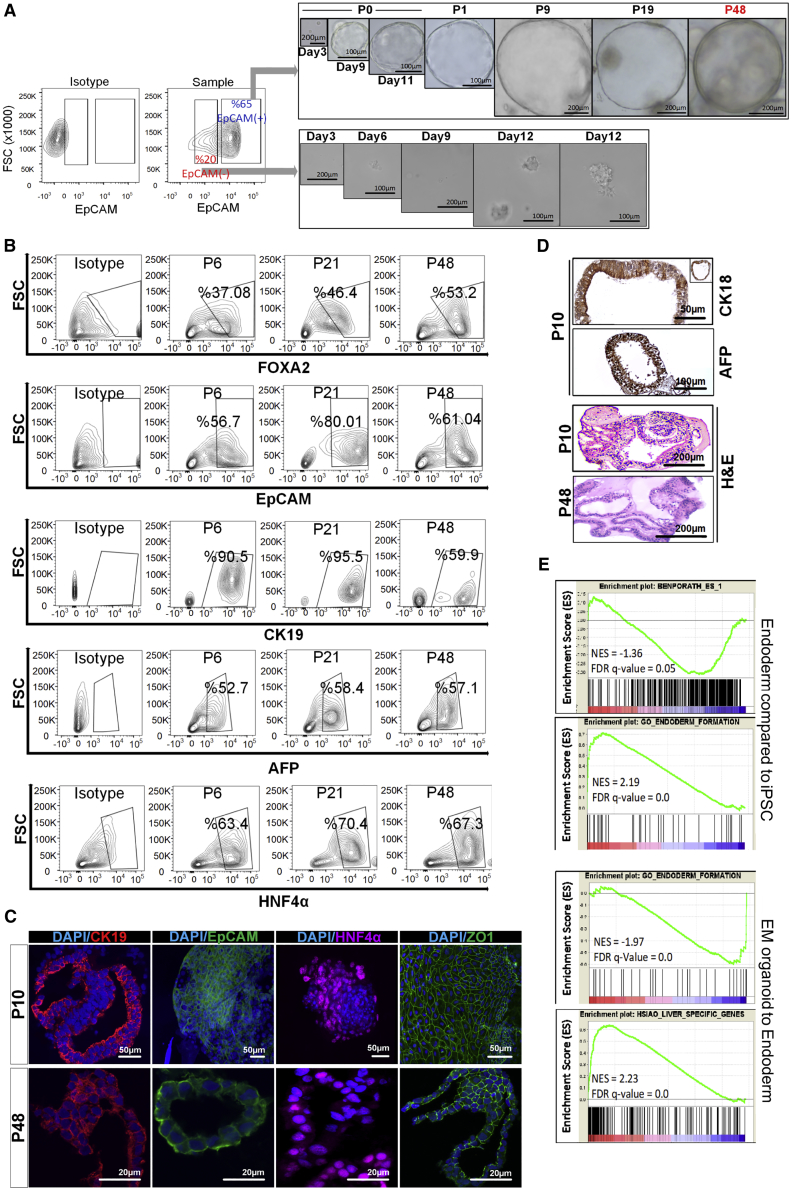
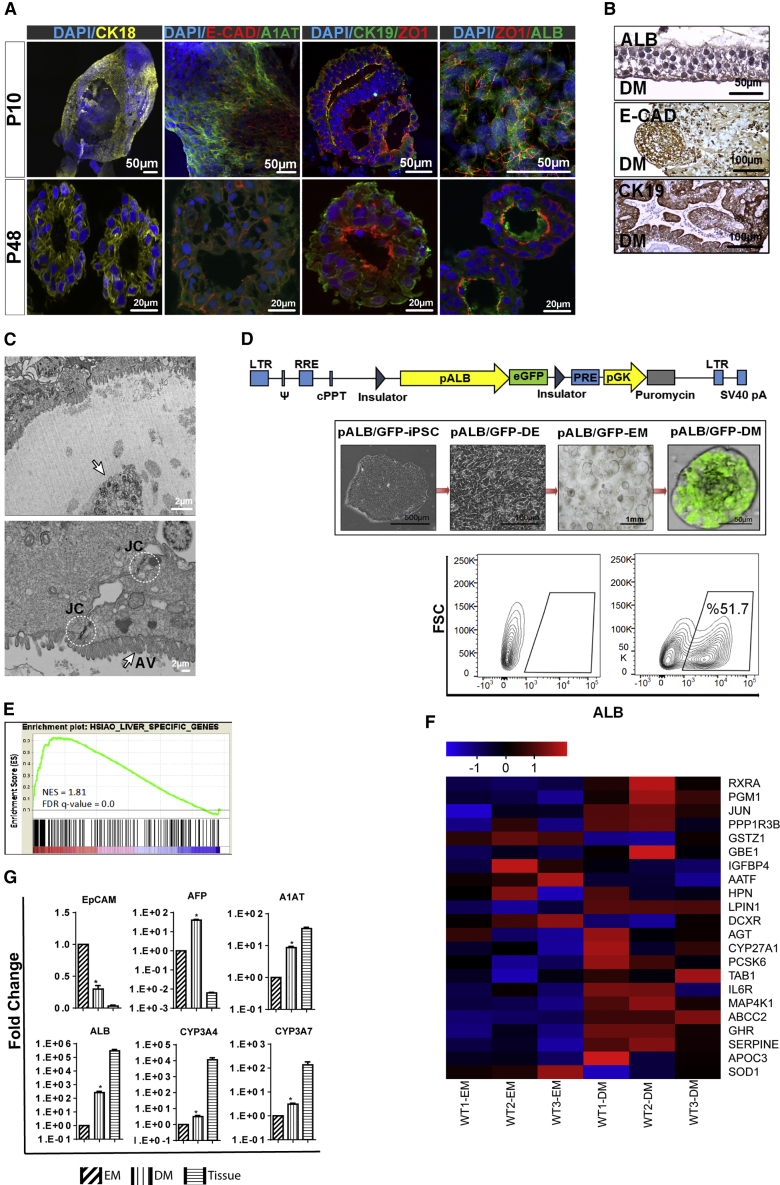
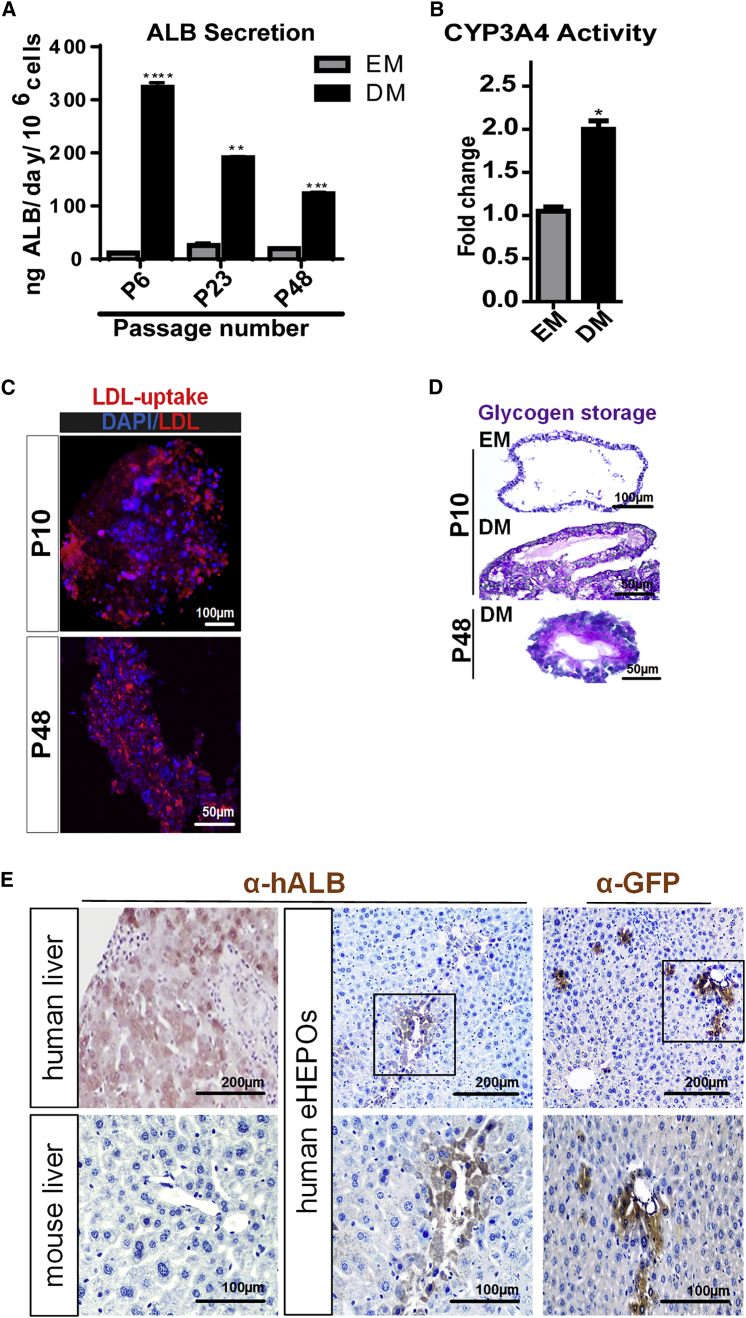
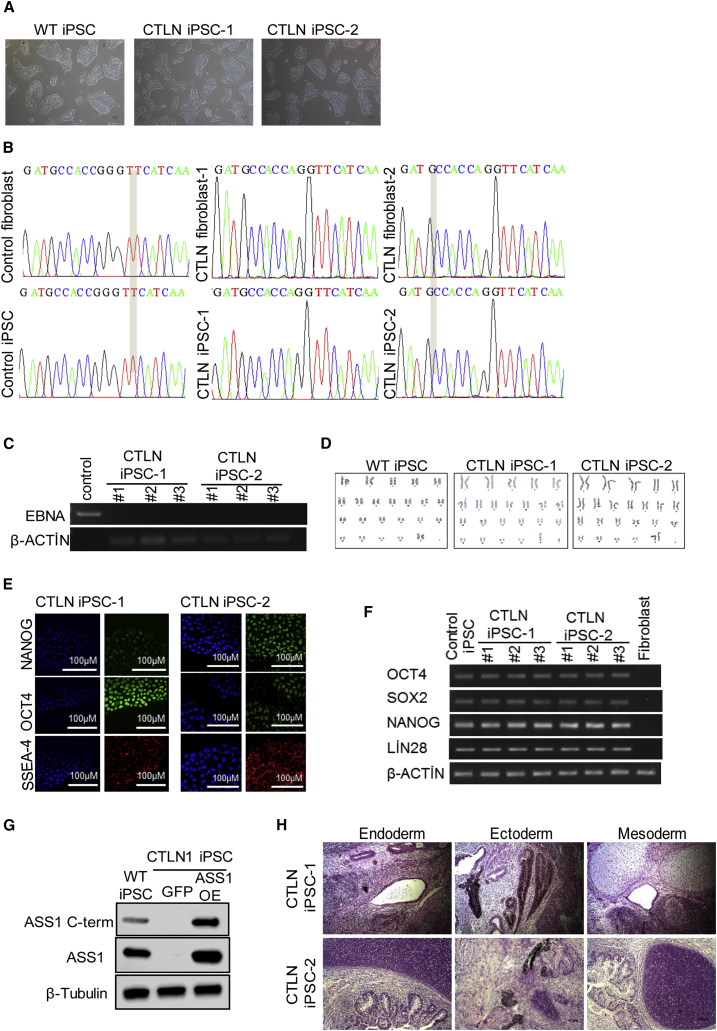
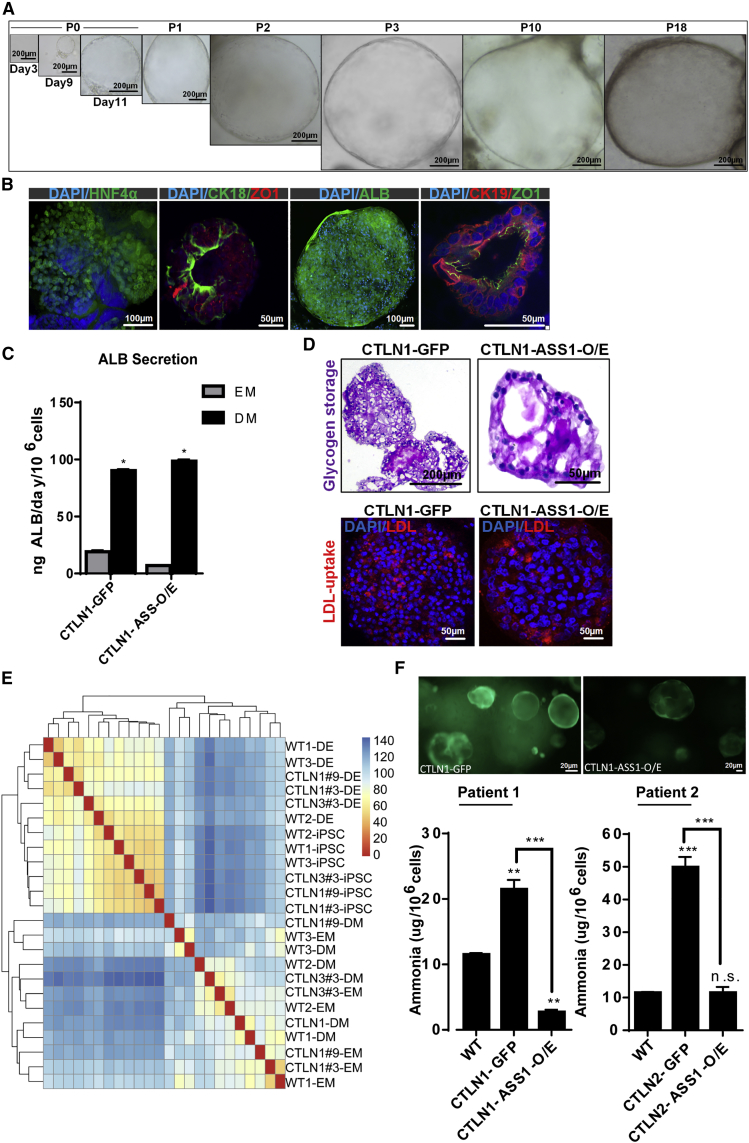
Organoid technologies have become a powerful emerging tool to model liver diseases, for drug screening, and for personalized treatments. These applications are, however, limited in their capacity to generate functional hepatocytes in a reproducible and efficient manner. Here, we generated and characterized the hepatic organoid (eHEPO) culture system using human induced pluripotent stem cell (iPSC)-derived EpCAM-positive endodermal cells as an intermediate. eHEPOs can be produced within 2 weeks and expanded long term (>16 months) without any loss of differentiation capacity to mature hepatocytes. Starting from patient-specific iPSCs, we modeled citrullinemia type 1, a urea cycle disorder caused by mutations in the argininosuccinate synthetase (ASS1) enzyme. The disease-related ammonia accumulation phenotype in eHEPOs could be reversed by the overexpression of the wild-type ASS1 gene, which also indicated that this model is amenable to genetic manipulation. Thus, eHEPOs are excellent unlimited cell sources to generate functional hepatic organoids in a fast and efficient manner.
Keywords: 3D organoid; EpCAM; citrullinemia; disease modelling; hepatocyte; hepatocyte differentiation; iPSC; liver.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Barker N., Huch M., Kujala P., van de Wetering M., Snippert H.J., van Es J.H., Sato T., Stange D.E., Begthel H., van den Born M. Lgr5 +ve stem cells drive self-renewal in the stomach and build long-lived gastric units in vitro. Cell Stem Cell. 2010;6:25–36. - PubMed
-
- Basak O., Beumer J., Wiebrands K., Seno H., van Oudenaarden A., Clevers H. Induced quiescence of Lgr5+ stem cells in intestinal organoids enables differentiation of hormone-producing enteroendocrine cells. Cell Stem Cell. 2017;20:177–190.e4. - PubMed
-
- Baxter M., Withey S., Harrison S., Segeritz C.-P., Zhang F., Atkinson-Dell R., Rowe C., Gerrard D.T., Sison-Young R., Jenkins R. Phenotypic and functional analyses show stem cell-derived hepatocyte-like cells better mimic fetal rather than adult hepatocytes. J. Hepatol. 2015;62:581–589. - PMC - PubMed
-
- Broutier L., Andersson-Rolf A., Hindley C.J., Boj S.F., Clevers H., Koo B.K., Huch M. Culture and establishment of self-renewing human and mouse adult liver and pancreas 3D organoids and their genetic manipulation. Nat. Protoc. 2016;11:1724–1743. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
