

Computational model demonstrates that Ndc80-associated proteins strengthen kinetochore-microtubule attachments in metaphase
- PMID: 31525284
- PMCID: PMC7641512
- DOI: 10.1002/cm.21562
Computational model demonstrates that Ndc80-associated proteins strengthen kinetochore-microtubule attachments in metaphase
Abstract
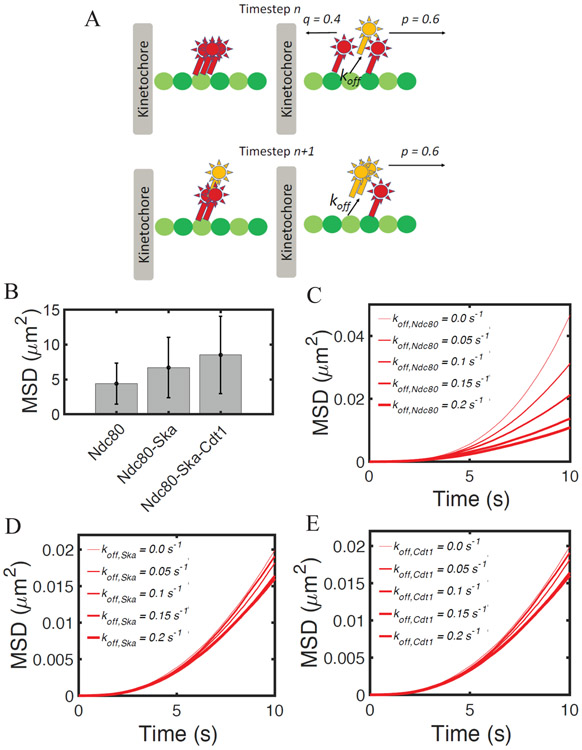
Chromosome segregation is mediated by spindle microtubules that attach to the kinetochore via dynamic protein complexes, such as Ndc80, Ska, Cdt1 and ch-TOG during mitotic metaphase. While experimental studies have previously shown that these proteins and protein complexes are all essential for maintaining a stable kinetochore-microtubule (kMT) interface, their exact roles in the mitotic metaphase remains elusive. In this study, we employed experimental and computational methods in order to characterize how these proteins can strengthen kMT attachments in both nonload-bearing and load-bearing conditions, typical of prometaphase and metaphase, respectively. Immunofluorescence staining of HeLa cells showed that the levels of Ska and Cdt1 significantly increased from prometaphase to metaphase, while levels of the Ndc80 complex remained unchanged. Our new computational model showed that by incorporating binding and unbinding of each protein complex coupled with a biased diffusion mechanism, the displacement of a possible complex formed by Ndc80-Ska-Cdt1 is significantly higher than that of Ndc80 alone or Ndc80-Ska. In addition, when we incorporate Ndc80/ch-TOG in the model, rupture force and time of attachment of the kMT interface increases. These results support the hypothesis that Ndc80-associated proteins strengthen kMT attachments, and that the interplay between kMT protein complexes in metaphase ensures stable attachments.
Keywords: Ndc80; kinetochore; microtubule; mitosis; modeling.
© 2019 Wiley Periodicals, Inc.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
