

Self-organization of modular network architecture by activity-dependent neuronal migration and outgrowth
- PMID: 31526478
- PMCID: PMC6783273
- DOI: 10.7554/eLife.47996
Self-organization of modular network architecture by activity-dependent neuronal migration and outgrowth
Abstract
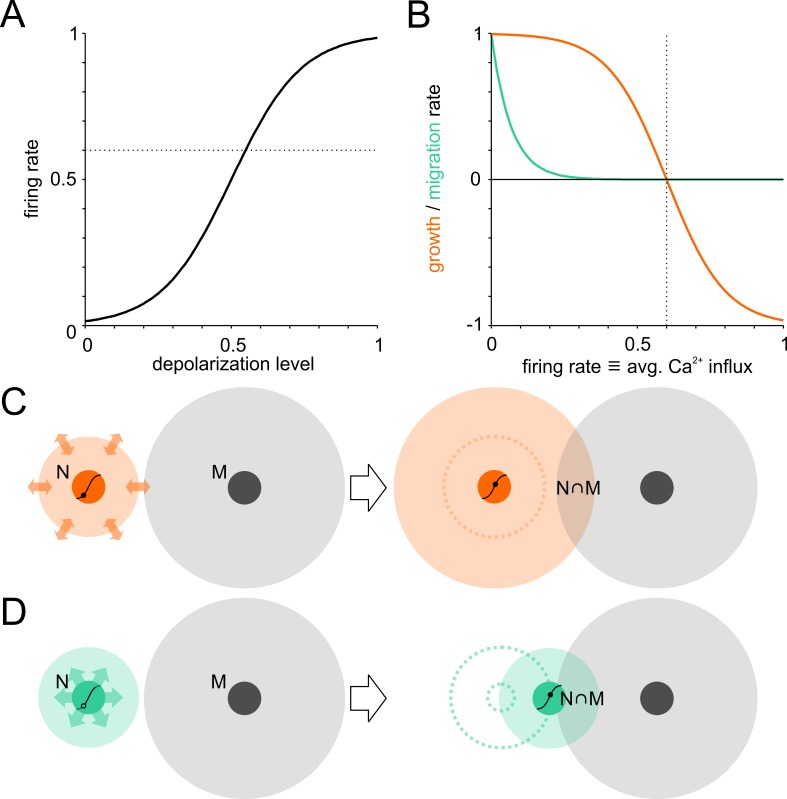
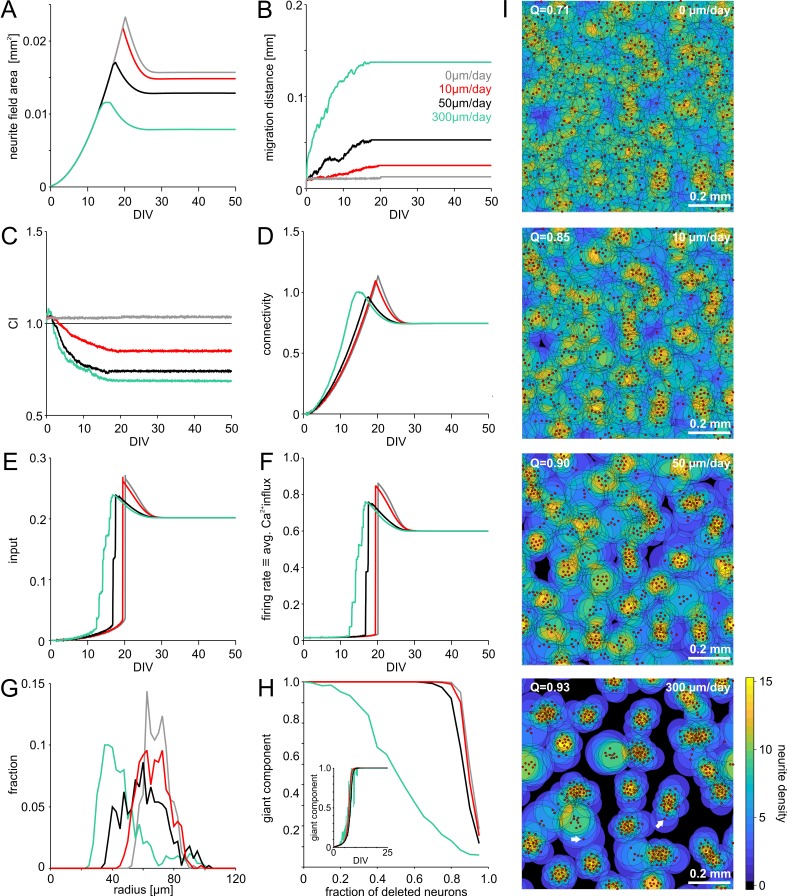
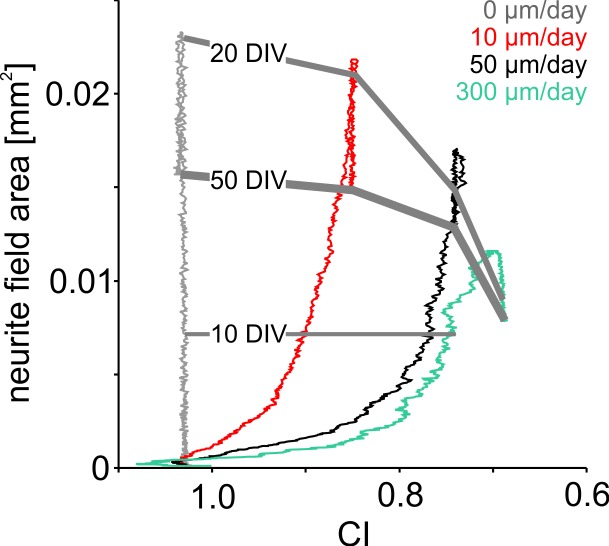
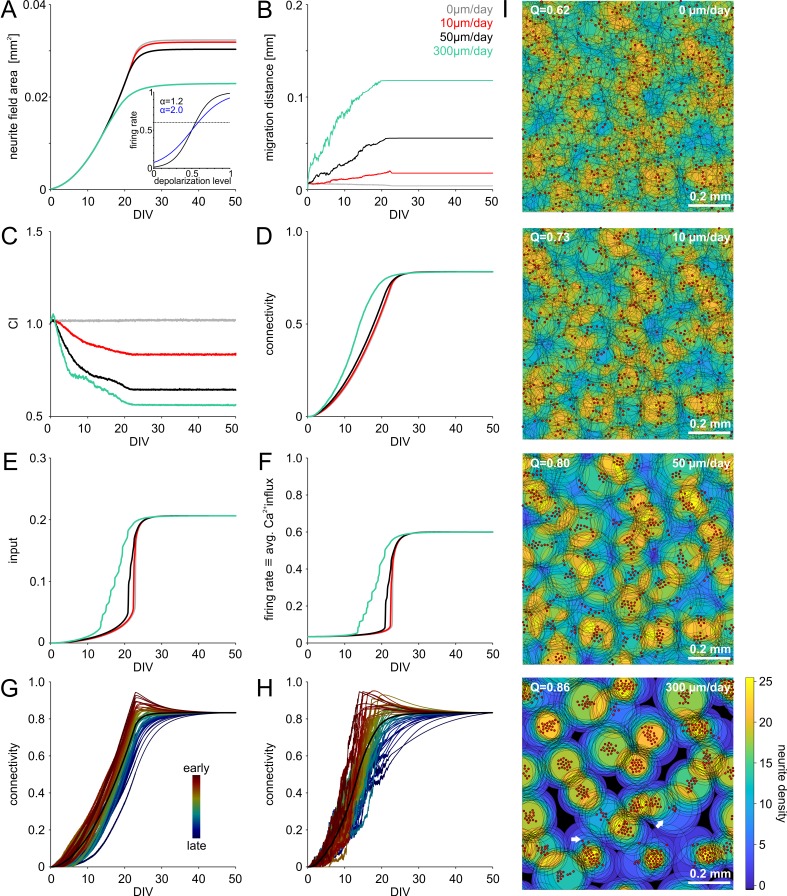
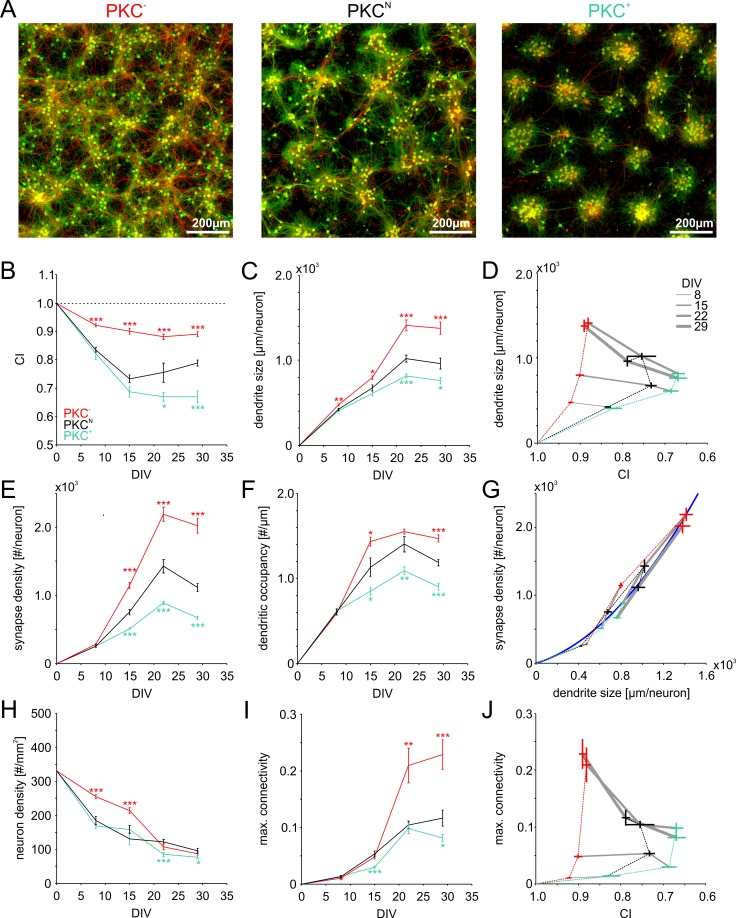
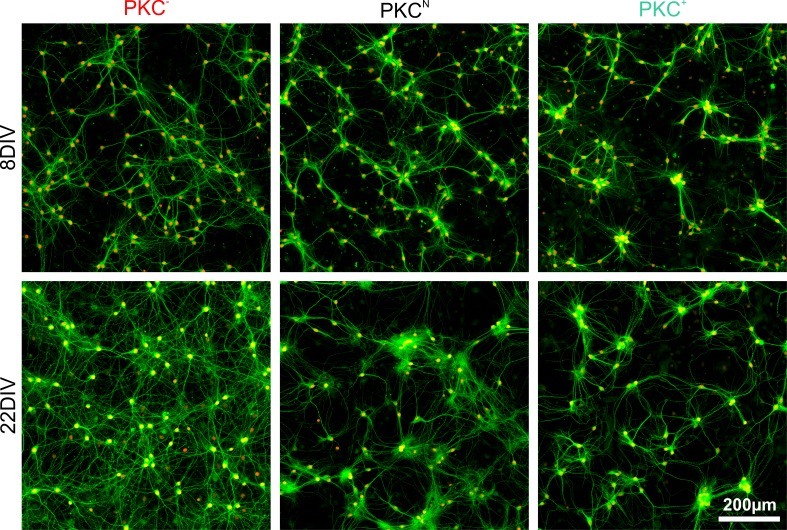
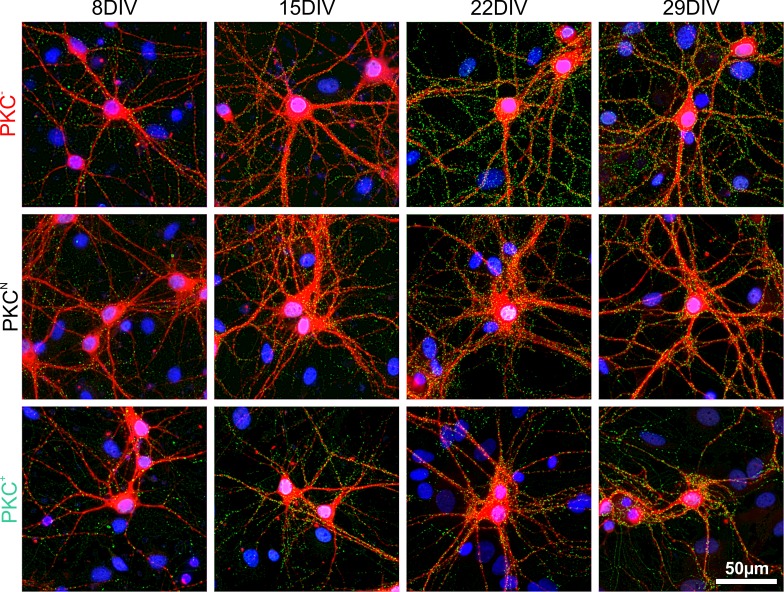
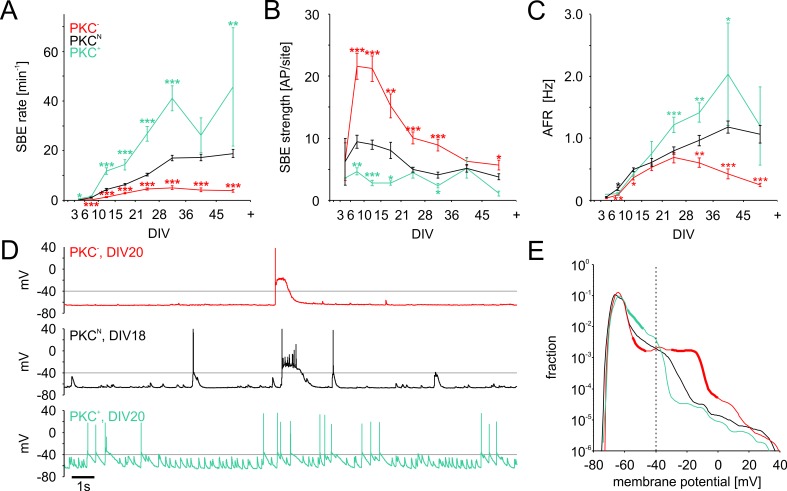
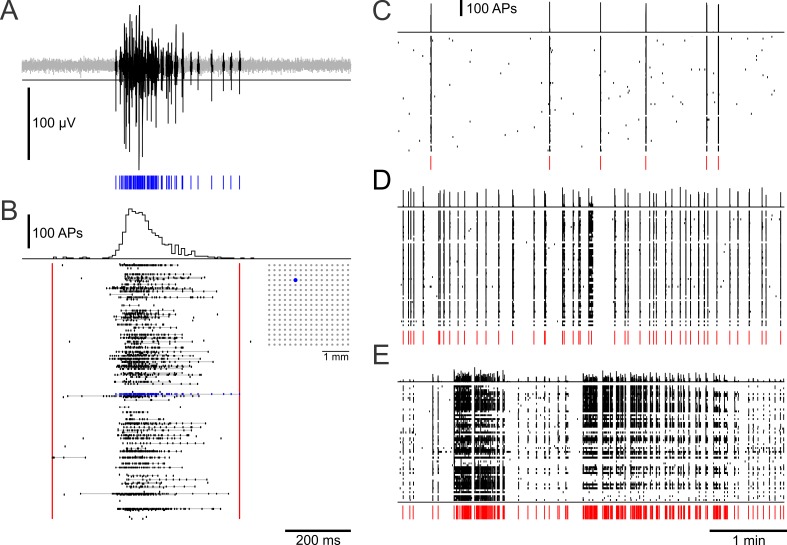
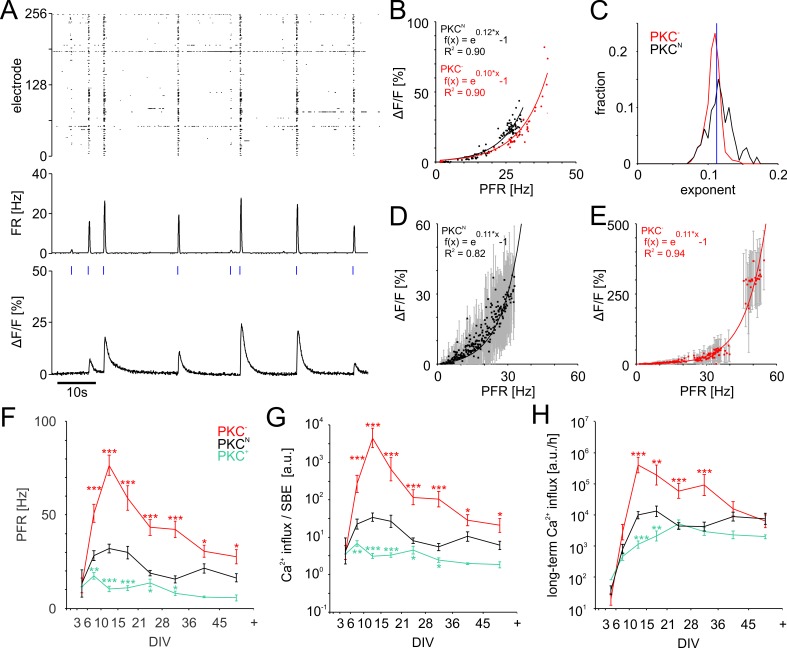
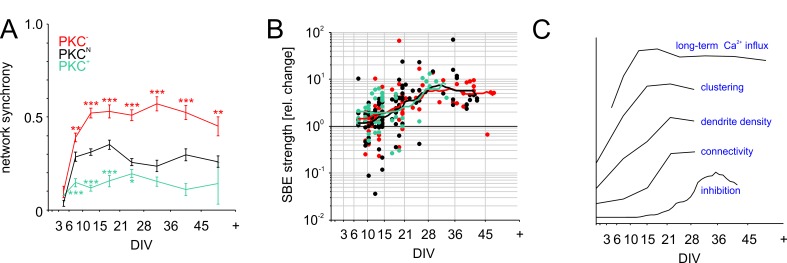
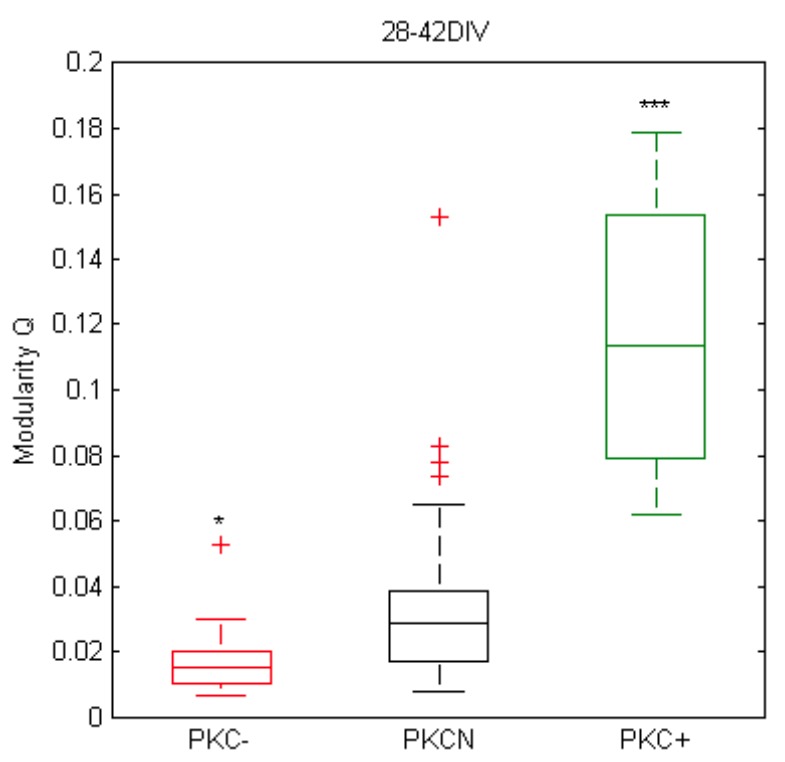
The spatial distribution of neurons and activity-dependent neurite outgrowth shape long-range interaction, recurrent local connectivity and the modularity in neuronal networks. We investigated how this mesoscale architecture develops by interaction of neurite outgrowth, cell migration and activity in cultured networks of rat cortical neurons and show that simple rules can explain variations of network modularity. In contrast to theoretical studies on activity-dependent outgrowth but consistent with predictions for modular networks, spontaneous activity and the rate of synchronized bursts increased with clustering, whereas peak firing rates in bursts increased in highly interconnected homogeneous networks. As Ca2+ influx increased exponentially with increasing network recruitment during bursts, its modulation was highly correlated to peak firing rates. During network maturation, long-term estimates of Ca2+ influx showed convergence, even for highly different mesoscale architectures, neurite extent, connectivity, modularity and average activity levels, indicating homeostatic regulation towards a common set-point of Ca2+ influx.
Keywords: activity-dependent development; homeostatic regulation; network structure; neurite outgrowth; neuronal migration; neuroscience; protein kinase C; rat.
© 2019, Okujeni and Egert.
Conflict of interest statement
SO, UE No competing interests declared
Figures
Similar articles
-
Mesoscale Architecture Shapes Initiation and Richness of Spontaneous Network Activity.J Neurosci. 2017 Apr 5;37(14):3972-3987. doi: 10.1523/JNEUROSCI.2552-16.2017. Epub 2017 Mar 14. J Neurosci. 2017. PMID: 28292833 Free PMC article.
-
Long-term characterization of firing dynamics of spontaneous bursts in cultured neural networks.IEEE Trans Biomed Eng. 2004 Nov;51(11):2051-62. doi: 10.1109/TBME.2004.827936. IEEE Trans Biomed Eng. 2004. PMID: 15536907
-
Structural Modularity Tunes Mesoscale Criticality in Biological Neuronal Networks.J Neurosci. 2023 Apr 5;43(14):2515-2526. doi: 10.1523/JNEUROSCI.1420-22.2023. Epub 2023 Mar 3. J Neurosci. 2023. PMID: 36868860 Free PMC article.
-
Self-organization and neuronal avalanches in networks of dissociated cortical neurons.Neuroscience. 2008 Jun 2;153(4):1354-69. doi: 10.1016/j.neuroscience.2008.03.050. Epub 2008 Mar 29. Neuroscience. 2008. PMID: 18448256
-
Homeostatic presynaptic suppression of neuronal network bursts.J Neurophysiol. 2009 Apr;101(4):2077-88. doi: 10.1152/jn.91085.2008. Epub 2009 Feb 4. J Neurophysiol. 2009. PMID: 19193770
Cited by
-
Rich dynamics and functional organization on topographically designed neuronal networks in vitro.iScience. 2022 Nov 26;25(12):105680. doi: 10.1016/j.isci.2022.105680. eCollection 2022 Dec 22. iScience. 2022. PMID: 36567712 Free PMC article.
-
Progenitor division and cell autonomous neurosecretion are required for rod photoreceptor sublaminar positioning.Proc Natl Acad Sci U S A. 2023 Oct 17;120(42):e2308204120. doi: 10.1073/pnas.2308204120. Epub 2023 Oct 9. Proc Natl Acad Sci U S A. 2023. PMID: 37812728 Free PMC article.
-
Gabaergic Interneurons in Early Brain Development: Conducting and Orchestrated by Cortical Network Activity.Front Mol Neurosci. 2022 Jan 3;14:807969. doi: 10.3389/fnmol.2021.807969. eCollection 2021. Front Mol Neurosci. 2022. PMID: 35046773 Free PMC article.
-
Tracking axon initial segment plasticity using high-density microelectrode arrays: A computational study.Front Neuroinform. 2022 Oct 3;16:957255. doi: 10.3389/fninf.2022.957255. Front Neuroinform. 2022. PMID: 36221258 Free PMC article.
-
Modular architecture facilitates noise-driven control of synchrony in neuronal networks.Sci Adv. 2023 Aug 25;9(34):eade1755. doi: 10.1126/sciadv.ade1755. Epub 2023 Aug 25. Sci Adv. 2023. PMID: 37624893 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
