

Early ear neuronal development, but not olfactory or lens development, can proceed without SOX2
- PMID: 31526806
- PMCID: PMC6938654
- DOI: 10.1016/j.ydbio.2019.09.003
Early ear neuronal development, but not olfactory or lens development, can proceed without SOX2
Abstract
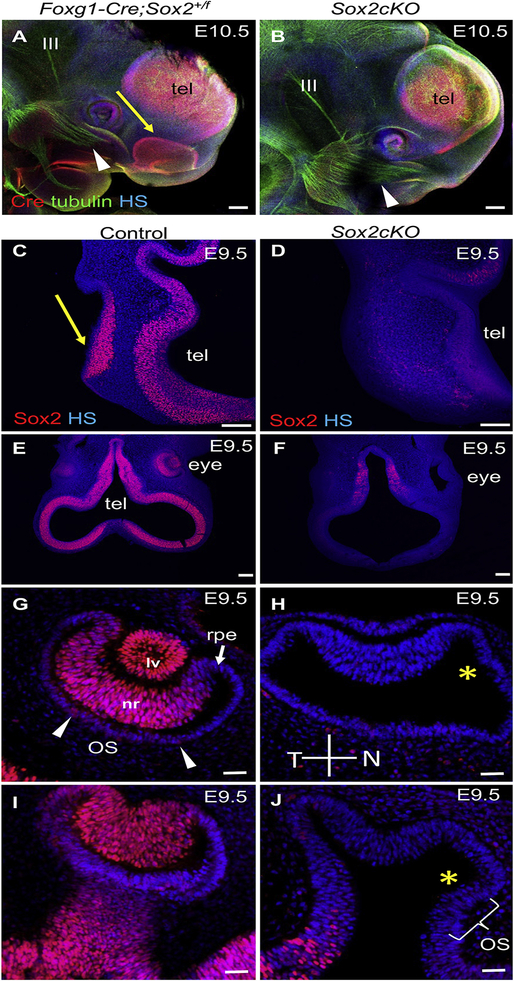
SOX2 is essential for maintaining neurosensory stem cell properties, although its involvement in the early neurosensory development of cranial placodes remains unclear. To address this, we used Foxg1-Cre to conditionally delete Sox2 during eye, ear, and olfactory placode development. Foxg1-Cre mediated early deletion of Sox2 eradicates all olfactory placode development, and disrupts retinal development and invagination of the lens placode. In contrast to the lens and olfactory placodes, the ear placode invaginates and delaminates NEUROD1 positive neurons. Furthermore, we show that SOX2 is not necessary for early ear neurogenesis, since the early inner ear ganglion is formed with near normal central projections to the hindbrain and peripheral projections to the undifferentiated sensory epithelia of E11.5-12.5 ears. However, later stages of ear neurosensory development, in particular, the late forming auditory system, critically depend on the presence of SOX2. Our data establish distinct differences for SOX2 requirements among placodal sensory organs with similarities between olfactory and lens but not ear placode development, consistent with the unique neurosensory development and molecular properties of the ear.
Keywords: Eye; Inner ear; Neuronal projections; Olfactory system; Placode development.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Competing interests
The authors have no competing interests to declare.
Figures











References
-
- Abello G, Khatri S, Radosevic M, Scotting PJ, Giraldez F, Alsina B, 2010. Independent regulation of Sox3 and Lmx1b by FGF and BMP signaling influences the neurogenic and non-neurogenic domains in the chick otic placode. Dev Biol 339, 166–178. - PubMed
-
- Barrionuevo F, Naumann A, Bagheri-Fam S, Speth V, Taketo MM, Scherer G, Neubuser A, 2008. Sox9 is required for invagination of the otic placode in mice. Dev Biol 317, 213–224. - PubMed
-
- Bhattacharyya S, Bailey AP, Bronner-Fraser M, Streit A, 2004. Segregation of lens and olfactory precursors from a common territory: cell sorting and reciprocity of Dlx5 and Pax6 expression. Dev Biol 271, 403–414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
