

Expression of a novel class of bacterial Ig-like proteins is required for IncHI plasmid conjugation
- PMID: 31527905
- PMCID: PMC6764697
- DOI: 10.1371/journal.pgen.1008399
Expression of a novel class of bacterial Ig-like proteins is required for IncHI plasmid conjugation
Abstract
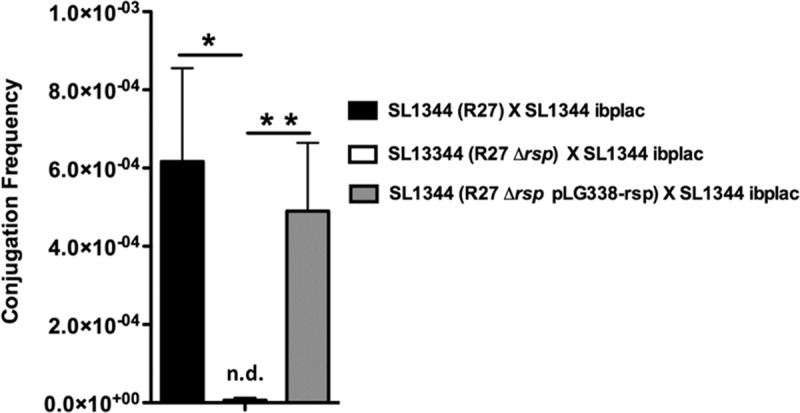
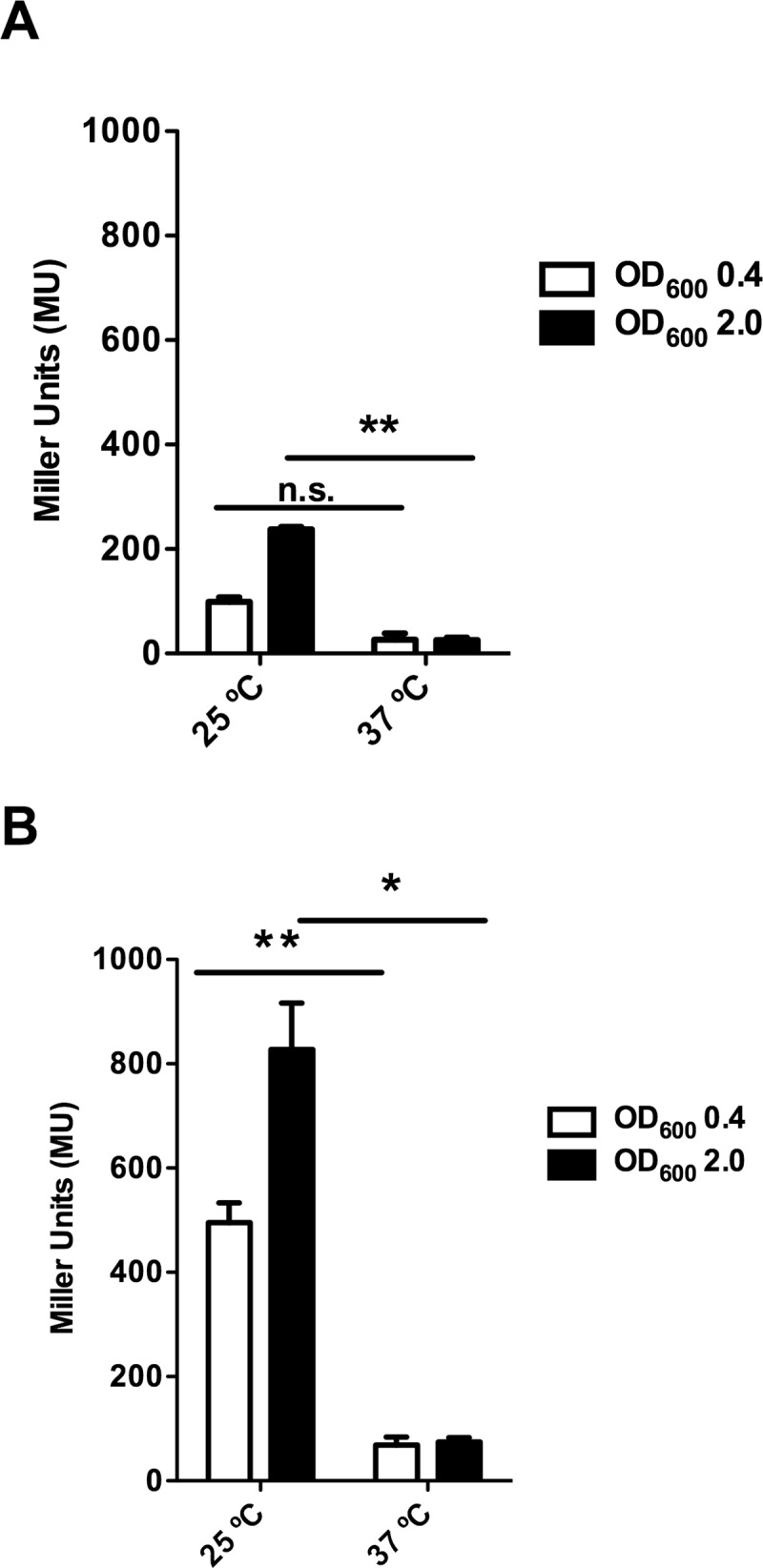
Antimicrobial resistance (AMR) is currently one of the most important challenges to the treatment of bacterial infections. A critical issue to combat AMR is to restrict its spread. In several instances, bacterial plasmids are involved in the global spread of AMR. Plasmids belonging to the incompatibility group (Inc)HI are widespread in Enterobacteriaceae and most of them express multiple antibiotic resistance determinants. They play a relevant role in the recent spread of colistin resistance. We present in this report novel findings regarding IncHI plasmid conjugation. Conjugative transfer in liquid medium of an IncHI plasmid requires expression of a plasmid-encoded, large-molecular-mass protein that contains an Ig-like domain. The protein, termed RSP, is encoded by a gene (ORF R0009) that maps in the Tra2 region of the IncHI1 R27 plasmid. The RSP protein is exported outside the cell by using the plasmid-encoded type IV secretion system that is also used for its transmission to new cells. Expression of the protein reduces cell motility and enables plasmid conjugation. Flagella are one of the cellular targets of the RSP protein. The RSP protein is required for a high rate of plasmid transfer in both flagellated and nonflagellated Salmonella cells. This effect suggests that RSP interacts with other cellular structures as well as with flagella. These unidentified interactions must facilitate mating pair formation and, hence, facilitate IncHI plasmid conjugation. Due to its location on the outer surfaces of the bacterial cell, targeting the RSP protein could be a means of controlling IncHI plasmid conjugation in natural environments or of combatting infections caused by AMR enterobacteria that harbor IncHI plasmids.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







References
-
- Rossolini GM, Mantengoli E, Docquier J-D, Musmanno RA, Coratza G. Epidemiology of infections caused by multiresistant gram-negatives: ESBLs, MBLs, panresistant strains. New Microbiol. 2007;30: 332–339. - PubMed
