

Contrasting Mixotrophic Lifestyles Reveal Different Ecological Niches in Two Closely Related Marine Protists
- PMID: 31529498
- PMCID: PMC7065223
- DOI: 10.1111/jpy.12920
Contrasting Mixotrophic Lifestyles Reveal Different Ecological Niches in Two Closely Related Marine Protists
Abstract
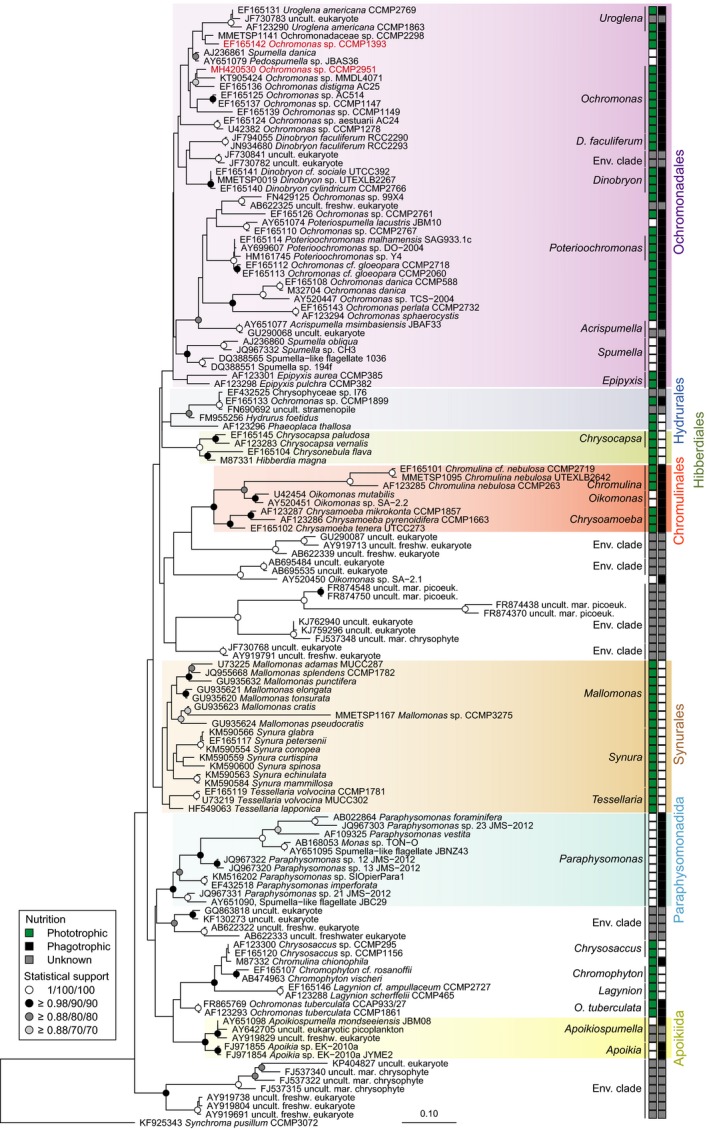
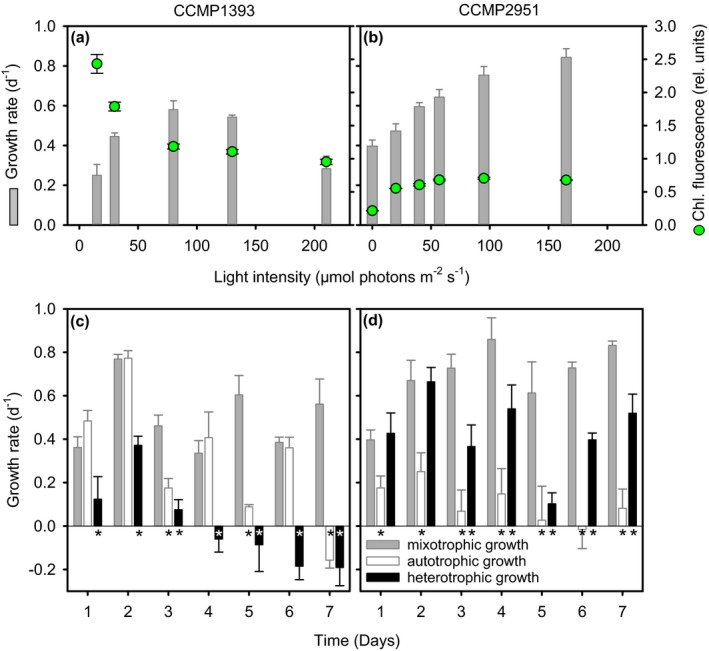
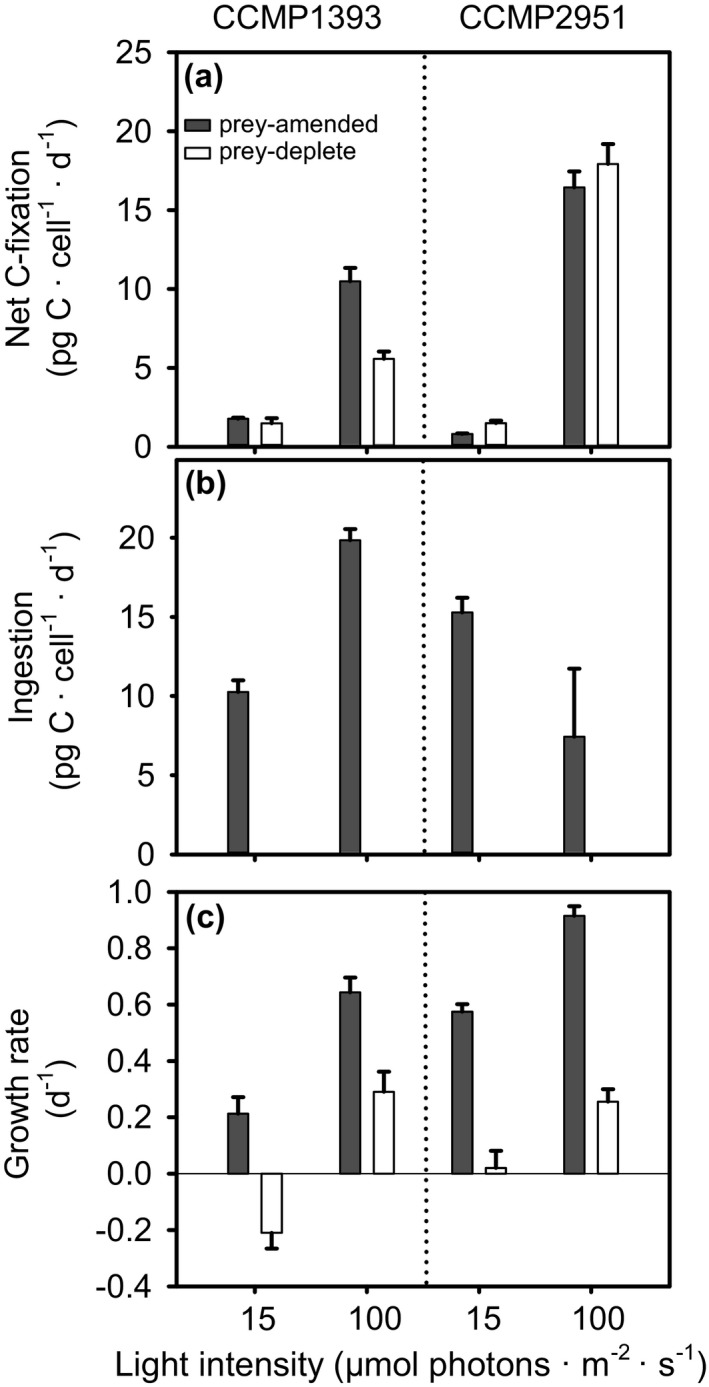
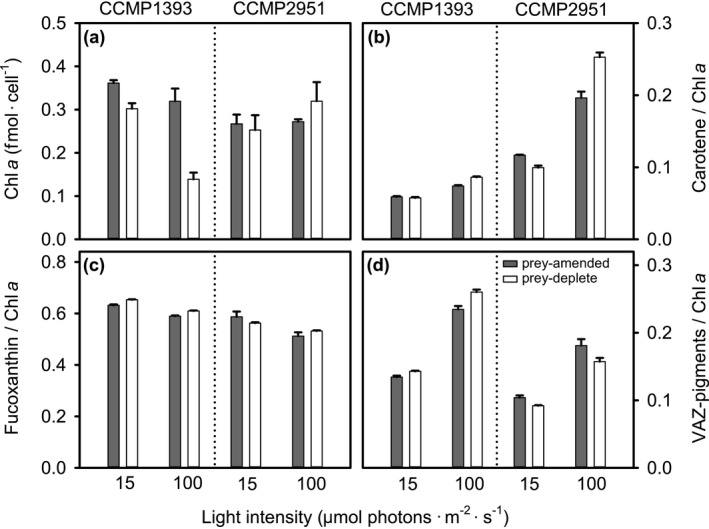
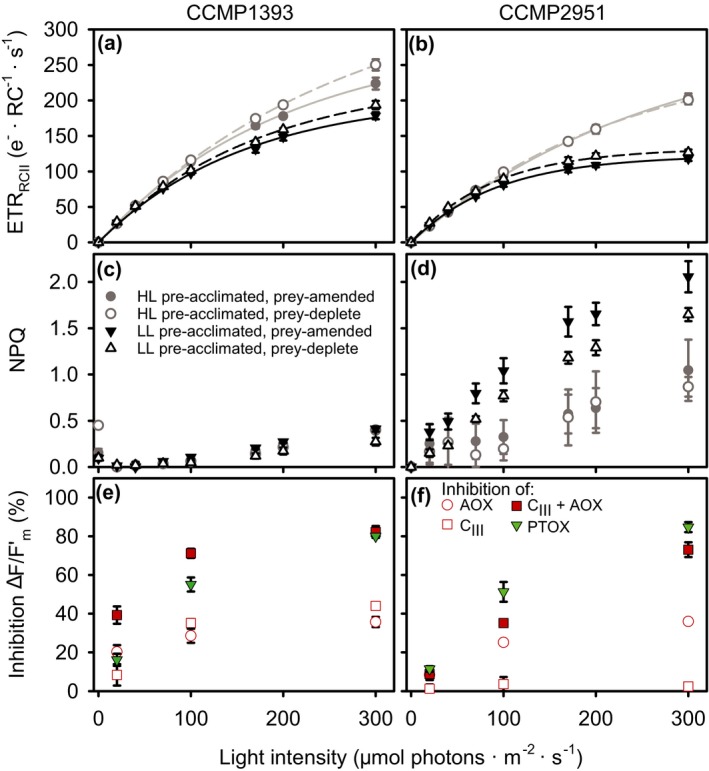
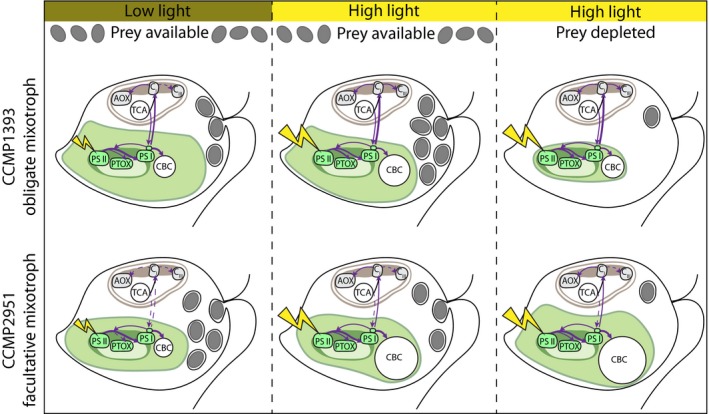
Many marine microbial eukaryotes combine photosynthetic with phagotrophic nutrition, but incomplete understanding of such mixotrophic protists, their functional diversity, and underlying physiological mechanisms limits the assessment and modeling of their roles in present and future ocean ecosystems. We developed an experimental system to study responses of mixotrophic protists to availability of living prey and light, and used it to characterize contrasting physiological strategies in two stramenopiles in the genus Ochromonas. We show that oceanic isolate CCMP1393 is an obligate mixotroph, requiring both light and prey as complementary resources. Interdependence of photosynthesis and heterotrophy in CCMP1393 comprises a significant role of mitochondrial respiration in photosynthetic electron transport. In contrast, coastal isolate CCMP2951 is a facultative mixotroph that can substitute photosynthesis by phagotrophy and hence grow purely heterotrophically in darkness. In contrast to CCMP1393, CCMP2951 also exhibits a marked photoprotection response that integrates non-photochemical quenching and mitochondrial respiration as electron sink for photosynthetically produced reducing equivalents. Facultative mixotrophs similar to CCMP2951 might be well adapted to variable environments, while obligate mixotrophs similar to CCMP1393 appear capable of resource efficient growth in oligotrophic ocean environments. Thus, the responses of these phylogenetically close protists to the availability of different resources reveals niche differentiation that influences impacts in food webs and leads to opposing carbon cycle roles.
Keywords: chrysophytes; microbial food web; mixotrophy; phagotrophy; phytoplankton.
© 2019 The Authors. Journal of Phycology published by Wiley Periodicals, Inc. on behalf of Phycological Society of America.
Figures
Similar articles
-
Variable responses to ocean acidification among mixotrophic protists with different lifestyles.ISME Commun. 2025 Apr 18;5(1):ycaf064. doi: 10.1093/ismeco/ycaf064. eCollection 2025 Jan. ISME Commun. 2025. PMID: 40391242 Free PMC article.
-
Mixotrophy in nanoflagellates across environmental gradients in the ocean.Proc Natl Acad Sci U S A. 2019 Mar 26;116(13):6211-6220. doi: 10.1073/pnas.1814860116. Epub 2019 Feb 13. Proc Natl Acad Sci U S A. 2019. PMID: 30760589 Free PMC article.
-
Modeling the metabolic evolution of mixotrophic phytoplankton in response to rising ocean surface temperatures.BMC Ecol Evol. 2022 Nov 18;22(1):136. doi: 10.1186/s12862-022-02092-9. BMC Ecol Evol. 2022. PMID: 36401160 Free PMC article.
-
Alternative nutritional strategies in protists: symposium introduction and a review of freshwater protists that combine photosynthesis and heterotrophy.J Eukaryot Microbiol. 2011 May-Jun;58(3):181-4. doi: 10.1111/j.1550-7408.2011.00543.x. Epub 2011 Apr 7. J Eukaryot Microbiol. 2011. PMID: 21477096 Review.
-
Mixotrophy everywhere on land and in water: the grand écart hypothesis.Ecol Lett. 2017 Feb;20(2):246-263. doi: 10.1111/ele.12714. Epub 2016 Dec 28. Ecol Lett. 2017. PMID: 28032461 Review.
Cited by
-
Broad phylogenetic and functional diversity among mixotrophic consumers of Prochlorococcus.ISME J. 2022 Jun;16(6):1557-1569. doi: 10.1038/s41396-022-01204-z. Epub 2022 Feb 10. ISME J. 2022. PMID: 35145244 Free PMC article.
-
Mixoplankton and mixotrophy: future research priorities.J Plankton Res. 2023 Jun 9;45(4):576-596. doi: 10.1093/plankt/fbad020. eCollection 2023 Jul-Aug. J Plankton Res. 2023. PMID: 37483910 Free PMC article. Review.
-
A multifunctional organelle coordinates phagocytosis and chlorophagy in a marine eukaryote phytoplankton Scyphosphaera apsteinii.New Phytol. 2025 May;246(3):1096-1112. doi: 10.1111/nph.20388. Epub 2025 Mar 4. New Phytol. 2025. PMID: 40035416 Free PMC article.
-
Integrated overview of stramenopile ecology, taxonomy, and heterotrophic origin.ISME J. 2024 Jan 8;18(1):wrae150. doi: 10.1093/ismejo/wrae150. ISME J. 2024. PMID: 39077993 Free PMC article. Review.
-
Light-dependent niche differentiation in two mixotrophic bacterivores.Environ Microbiol Rep. 2022 Aug;14(4):530-537. doi: 10.1111/1758-2229.13071. Epub 2022 May 4. Environ Microbiol Rep. 2022. PMID: 35509225 Free PMC article.
References
-
- Andersen, K. H. , Aksnes, D. L. , Berge, T. , Fiksen, Ø. & Visser, A. 2015. Modelling emergent trophic strategies in plankton. J. Plankton Res. 37:862–8.
-
- Andersson, A. , Falk, S. , Samuelsson, G. & Hagström, Å. 1989. Nutritional characteristics of a mixotrophic nanoflagellate, Ochromonas sp. Microb. Ecol. 17:251–62. - PubMed
-
- Arsalane, W. , Rousseau, B. & Duval, J. 1994. Influence of the pool size of the xanthophyll cycle on the effects of light stress in a diatom: Competition between photoprotection and photoinhibition. Photochem. Photobiol. 60:237–43.
-
- Azam, F. , Fenchel, T. , Field, J. G. , Gray, J. S. , Meyerreil, L. A. & Thingstad, F. 1983. The ecological role of water‐column microbes in the sea. Mar. Ecol. Prog. Ser. 10:257–63.
Publication types
MeSH terms
Associated data
- Actions
LinkOut - more resources
Full Text Sources
