

Rare Human Missense Variants can affect the Function of Disease-Relevant Proteins by Loss and Gain of Peroxisomal Targeting Motifs
- PMID: 31533369
- PMCID: PMC6770196
- DOI: 10.3390/ijms20184609
Rare Human Missense Variants can affect the Function of Disease-Relevant Proteins by Loss and Gain of Peroxisomal Targeting Motifs
Abstract
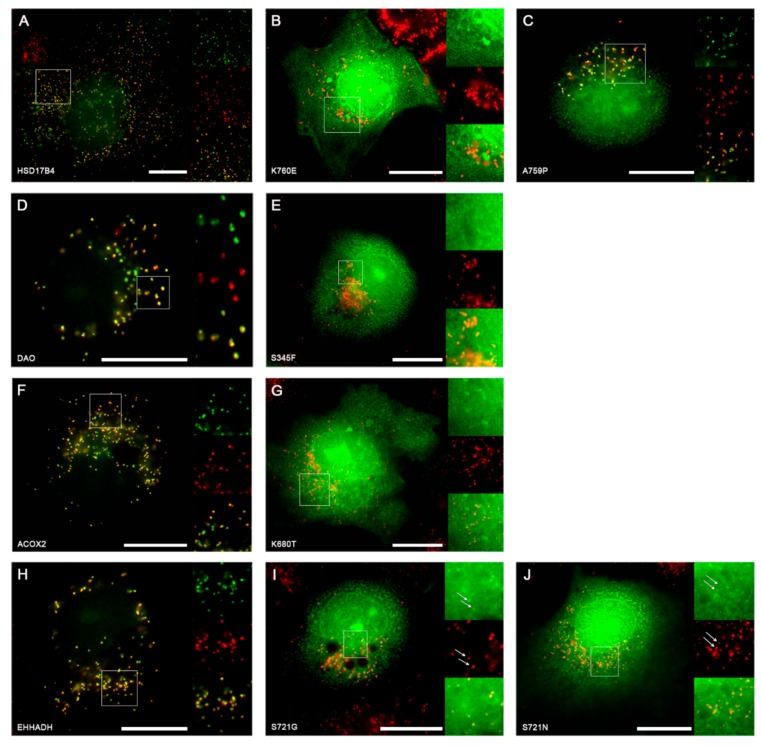
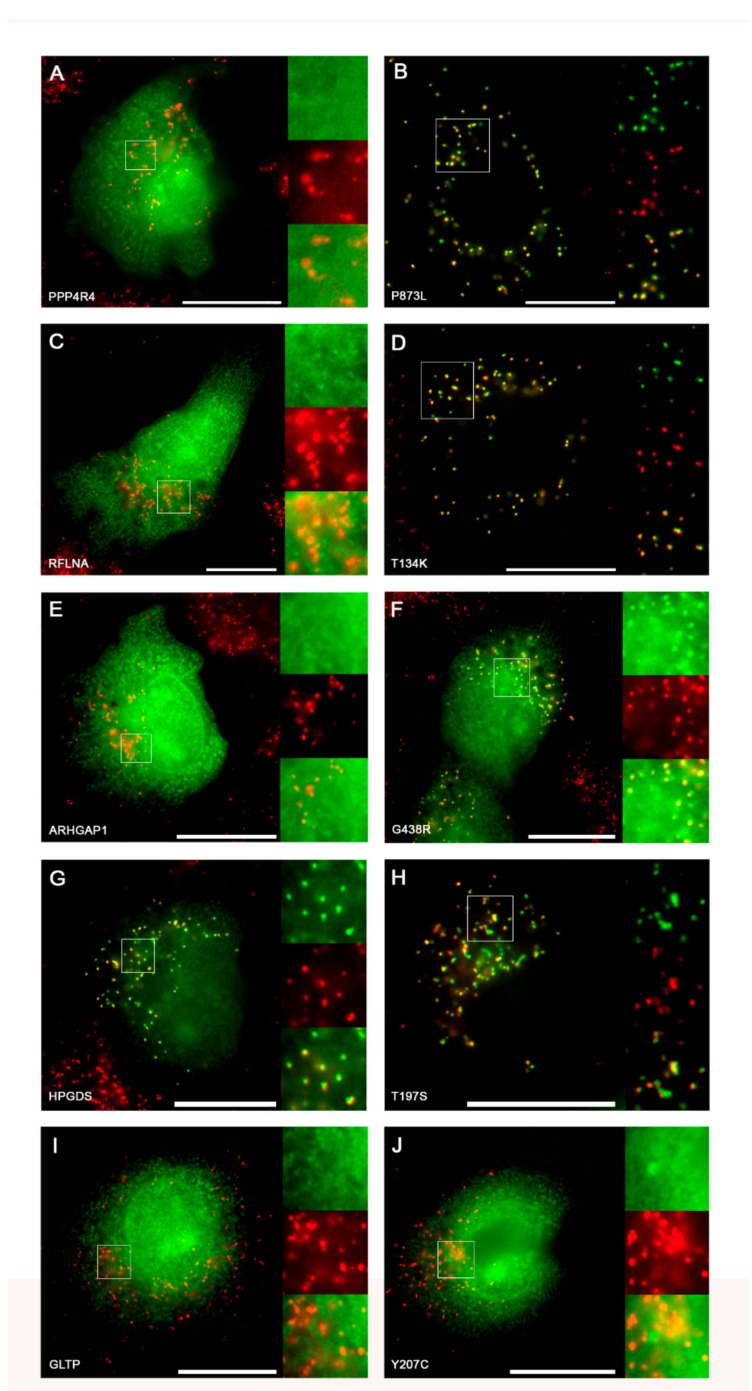
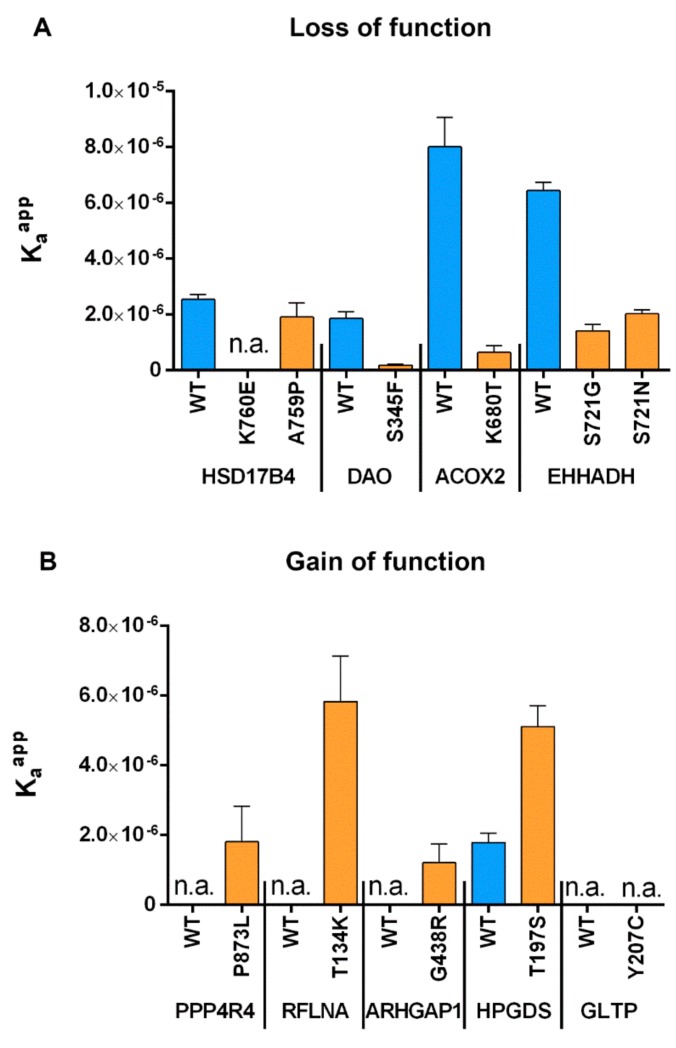
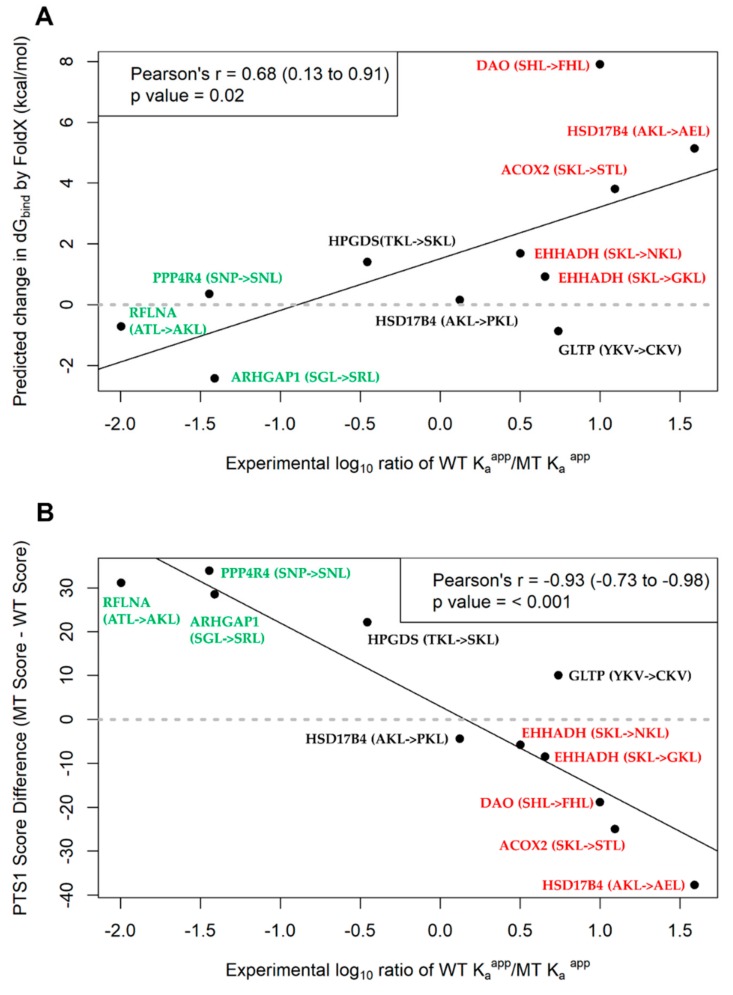
Single nucleotide variants (SNVs) resulting in amino acid substitutions (i.e., missense variants) can affect protein localization by changing or creating new targeting signals. Here, we studied the potential of naturally occurring SNVs from the Genome Aggregation Database (gnomAD) to result in the loss of an existing peroxisomal targeting signal 1 (PTS1) or gain of a novel PTS1 leading to mistargeting of cytosolic proteins to peroxisomes. Filtering down from 32,985 SNVs resulting in missense mutations within the C-terminal tripeptide of 23,064 human proteins, based on gene annotation data and computational prediction, we selected six SNVs for experimental testing of loss of function (LoF) of the PTS1 motif and five SNVs in cytosolic proteins for gain in PTS1-mediated peroxisome import (GoF). Experimental verification by immunofluorescence microscopy for subcellular localization and FRET affinity measurements for interaction with the receptor PEX5 demonstrated that five of the six predicted LoF SNVs resulted in loss of the PTS1 motif while three of five predicted GoF SNVs resulted in de novo PTS1 generation. Overall, we showed that a complementary approach incorporating bioinformatics methods and experimental testing was successful in identifying SNVs capable of altering peroxisome protein import, which may have implications in human disease.
Keywords: PEX5; disease; gnomAD; missense variant; mistargeting; peroxisomal targeting signal 1; peroxisome; protein transport.
Conflict of interest statement
The authors declare no conflict of interests.
Figures


Similar articles
-
A missense allele of PEX5 is responsible for the defective import of PTS2 cargo proteins into peroxisomes.Hum Genet. 2021 Apr;140(4):649-666. doi: 10.1007/s00439-020-02238-z. Epub 2021 Jan 2. Hum Genet. 2021. PMID: 33389129
-
Peroxisomal targeting signal receptor Pex5p interacts with cargoes and import machinery components in a spatiotemporally differentiated manner: conserved Pex5p WXXXF/Y motifs are critical for matrix protein import.Mol Cell Biol. 2002 Mar;22(6):1639-55. doi: 10.1128/MCB.22.6.1639-1655.2002. Mol Cell Biol. 2002. PMID: 11865044 Free PMC article.
-
Redox-regulated cargo binding and release by the peroxisomal targeting signal receptor, Pex5.J Biol Chem. 2013 Sep 20;288(38):27220-27231. doi: 10.1074/jbc.M113.492694. Epub 2013 Jul 31. J Biol Chem. 2013. PMID: 23902771 Free PMC article.
-
Characterization, prediction and evolution of plant peroxisomal targeting signals type 1 (PTS1s).Biochim Biophys Acta. 2016 May;1863(5):790-803. doi: 10.1016/j.bbamcr.2016.01.001. Epub 2016 Jan 6. Biochim Biophys Acta. 2016. PMID: 26772785 Review.
-
Peroxisome targeting signal 1: is it really a simple tripeptide?Biochim Biophys Acta. 2006 Dec;1763(12):1565-73. doi: 10.1016/j.bbamcr.2006.08.022. Epub 2006 Aug 24. Biochim Biophys Acta. 2006. PMID: 17007944 Review.
Cited by
-
Molecular Role of Asn680Ser and Asp37Glu Missense Variants in Saudi Women with Female Infertility and Polycystic Ovarian Syndrome.Curr Issues Mol Biol. 2023 Jun 29;45(7):5494-5514. doi: 10.3390/cimb45070348. Curr Issues Mol Biol. 2023. PMID: 37504264 Free PMC article.
-
Insights Into the Peroxisomal Protein Inventory of Zebrafish.Front Physiol. 2022 Feb 28;13:822509. doi: 10.3389/fphys.2022.822509. eCollection 2022. Front Physiol. 2022. PMID: 35295584 Free PMC article.
-
Studying the interaction between PEX5 and its full-length cargo proteins in living cells by a novel Försteŕs resonance energy transfer-based competition assay.Front Cell Dev Biol. 2022 Nov 3;10:1026388. doi: 10.3389/fcell.2022.1026388. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36407094 Free PMC article.
-
Signal Peptides: From Molecular Mechanisms to Applications in Protein and Vaccine Engineering.Biomolecules. 2025 Jun 18;15(6):897. doi: 10.3390/biom15060897. Biomolecules. 2025. PMID: 40563537 Free PMC article. Review.
References
-
- Stenson P.D., Mort M., Ball E.V., Shaw K., Phillips A., Cooper D.N. The Human Gene Mutation Database: Building a comprehensive mutation repository for clinical and molecular genetics, diagnostic testing and personalized genomic medicine. Hum. Genet. 2014;133:1–9. doi: 10.1007/s00439-013-1358-4. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
