

Recovirus NS1-2 Has Viroporin Activity That Induces Aberrant Cellular Calcium Signaling To Facilitate Virus Replication
- PMID: 31533997
- PMCID: PMC6751491
- DOI: 10.1128/mSphere.00506-19
Recovirus NS1-2 Has Viroporin Activity That Induces Aberrant Cellular Calcium Signaling To Facilitate Virus Replication
Abstract
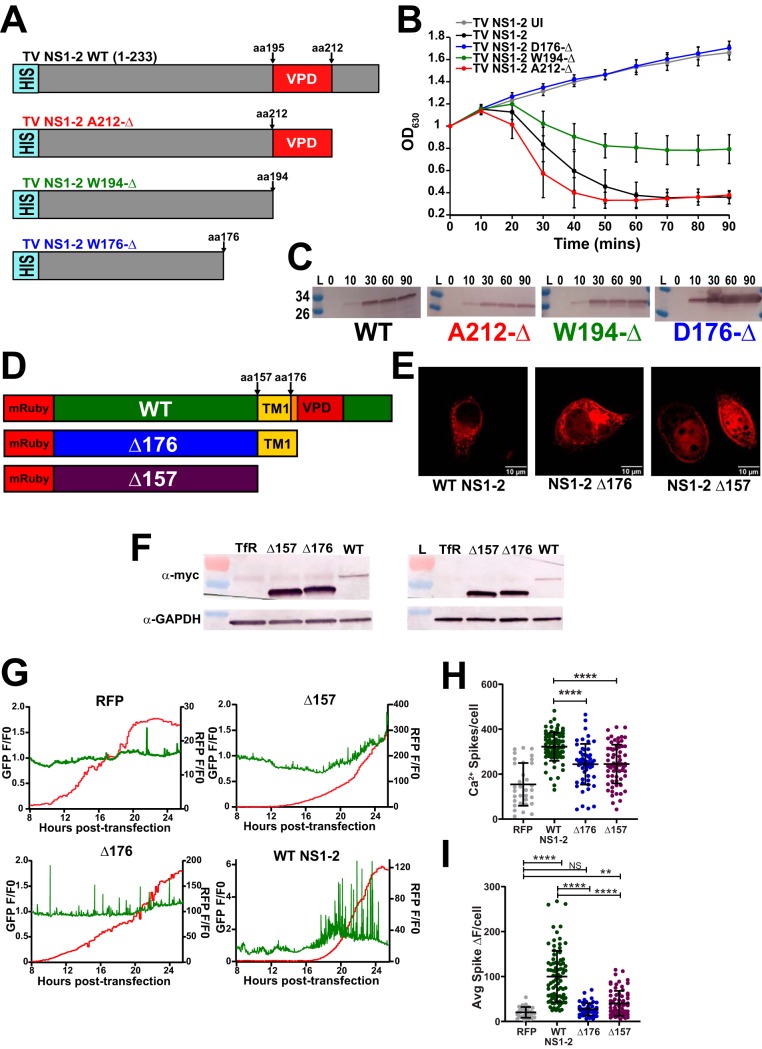
Enteric viruses in the Caliciviridae family cause acute gastroenteritis in humans and animals, but the cellular processes needed for virus replication and disease remain unknown. A common strategy among enteric viruses, including rotaviruses and enteroviruses, is to encode a viral ion channel (i.e., viroporin) that is targeted to the endoplasmic reticulum (ER) and disrupts host calcium (Ca2+) homeostasis. Previous reports have demonstrated genetic and functional similarities between the nonstructural proteins of caliciviruses and enteroviruses, including the calicivirus NS1-2 protein and the 2B viroporin of enteroviruses. However, it is unknown whether caliciviruses alter Ca2+ homeostasis for virus replication or whether the NS1-2 protein has viroporin activity like its enterovirus counterpart. To address these questions, we used Tulane virus (TV), a rhesus enteric calicivirus, to examine Ca2+ signaling during infection and determine whether NS1-2 has viroporin activity that disrupts Ca2+ homeostasis. We found that TV increases Ca2+ signaling during infection and that increased cytoplasmic Ca2+ levels are important for efficient replication. Further, TV NS1-2 localizes to the endoplasmic reticulum, the predominant intracellular Ca2+ store, and the NS2 region has characteristics of a viroporin domain (VPD). NS1-2 had viroporin activity in a classic bacterial functional assay and caused aberrant Ca2+ signaling when expressed in mammalian cells, but truncation of the VPD abrogated these activities. Together, our data provide new mechanistic insights into the function of the NS2 region of NS1-2 and support the premise that enteric viruses, including those within Caliciviridae, exploit host Ca2+ signaling to facilitate their replication.IMPORTANCE Tulane virus is one of many enteric caliciviruses that cause acute gastroenteritis and diarrheal disease. Globally, enteric caliciviruses affect both humans and animals and amass >65 billion dollars per year in treatment and health care-associated costs, thus imposing an enormous economic burden. Recent progress has resulted in several cultivation systems (B cells, enteroids, and zebrafish larvae) to study human noroviruses, but mechanistic insights into the viral factors and host pathways important for enteric calicivirus replication and infection are still largely lacking. Here, we used Tulane virus, a calicivirus that is biologically similar to human noroviruses and can be cultivated by conventional cell culture, to identify and functionally validate NS1-2 as an enteric calicivirus viroporin. Viroporin-mediated calcium signaling may be a broadly utilized pathway for enteric virus replication, and its existence within caliciviruses provides a novel approach to developing antivirals and comprehensive therapeutics for enteric calicivirus diarrheal disease outbreaks.
Keywords: GCaMP; calcium; calicivirus; viroporin.
Copyright © 2019 Strtak et al.
Figures






Similar articles
-
Rotavirus disrupts calcium homeostasis by NSP4 viroporin activity.mBio. 2010 Nov 30;1(5):e00265-10. doi: 10.1128/mBio.00265-10. mBio. 2010. PMID: 21151776 Free PMC article.
-
The Coxsackievirus and Adenovirus Receptor, a Required Host Factor for Recovirus Infection, Is a Putative Enteric Calicivirus Receptor.J Virol. 2019 Oct 29;93(22):e00869-19. doi: 10.1128/JVI.00869-19. Print 2019 Nov 15. J Virol. 2019. PMID: 31484750 Free PMC article.
-
Bat Caliciviruses and Human Noroviruses Are Antigenically Similar and Have Overlapping Histo-Blood Group Antigen Binding Profiles.mBio. 2018 May 22;9(3):e00869-18. doi: 10.1128/mBio.00869-18. mBio. 2018. PMID: 29789360 Free PMC article.
-
Pathophysiological Consequences of Calcium-Conducting Viroporins.Annu Rev Virol. 2015 Nov;2(1):473-96. doi: 10.1146/annurev-virology-100114-054846. Annu Rev Virol. 2015. PMID: 26958925 Free PMC article. Review.
-
Role of histo-blood group antigens in primate enteric calicivirus infections.World J Virol. 2014 Aug 12;3(3):18-21. doi: 10.5501/wjv.v3.i3.18. World J Virol. 2014. PMID: 25392814 Free PMC article. Review.
Cited by
-
Decoding mechanisms of diarrhea induction by enteric viruses.PLoS Pathog. 2024 Aug 8;20(8):e1012414. doi: 10.1371/journal.ppat.1012414. eCollection 2024 Aug. PLoS Pathog. 2024. PMID: 39116034 Free PMC article. Review. No abstract available.
-
The Feline Calicivirus Leader of the Capsid Protein Has the Functional Characteristics of a Viroporin.Viruses. 2022 Mar 18;14(3):635. doi: 10.3390/v14030635. Viruses. 2022. PMID: 35337042 Free PMC article.
-
Functions of Viroporins in the Viral Life Cycle and Their Regulation of Host Cell Responses.Front Immunol. 2022 Jun 2;13:890549. doi: 10.3389/fimmu.2022.890549. eCollection 2022. Front Immunol. 2022. PMID: 35720341 Free PMC article. Review.
-
Heterocyclic Inhibitors of Viroporins in the Design of Antiviral Compounds.Chem Heterocycl Compd (N Y). 2020;56(6):626-635. doi: 10.1007/s10593-020-02712-6. Epub 2020 Jul 17. Chem Heterocycl Compd (N Y). 2020. PMID: 32836315 Free PMC article.
-
Calcium Ions Signaling: Targets for Attack and Utilization by Viruses.Front Microbiol. 2022 Jul 4;13:889374. doi: 10.3389/fmicb.2022.889374. eCollection 2022. Front Microbiol. 2022. PMID: 35859744 Free PMC article. Review.
References
-
- Wobus CE, Cunha JB, Elftman M, Kolawole AO. 2016. Animal models of norovirus infection, p 397–415. In Svensson L, Desselberger U, Greenberg HB, Estes MK (ed), Viral gastroenteritis: molecular epidemiology and pathogenesis. Academic Press, Cambridge, MA.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
