

Stunted microbiota and opportunistic pathogen colonization in caesarean-section birth
- PMID: 31534227
- PMCID: PMC6894937
- DOI: 10.1038/s41586-019-1560-1
Stunted microbiota and opportunistic pathogen colonization in caesarean-section birth
Abstract
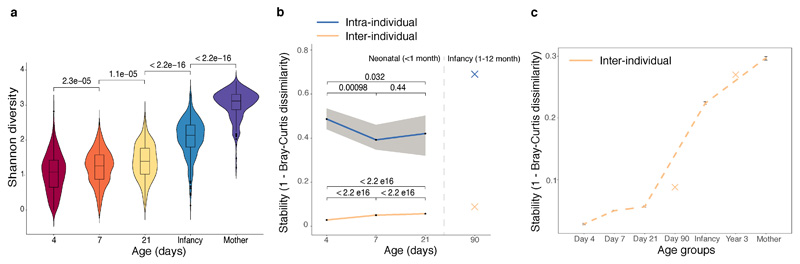
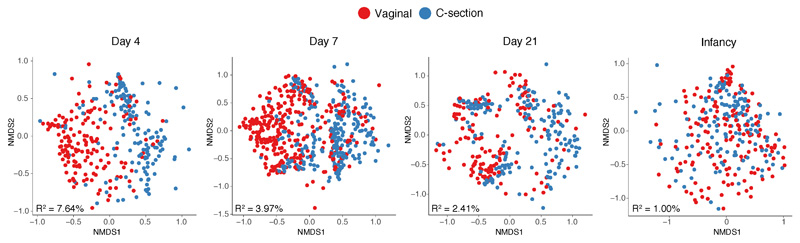
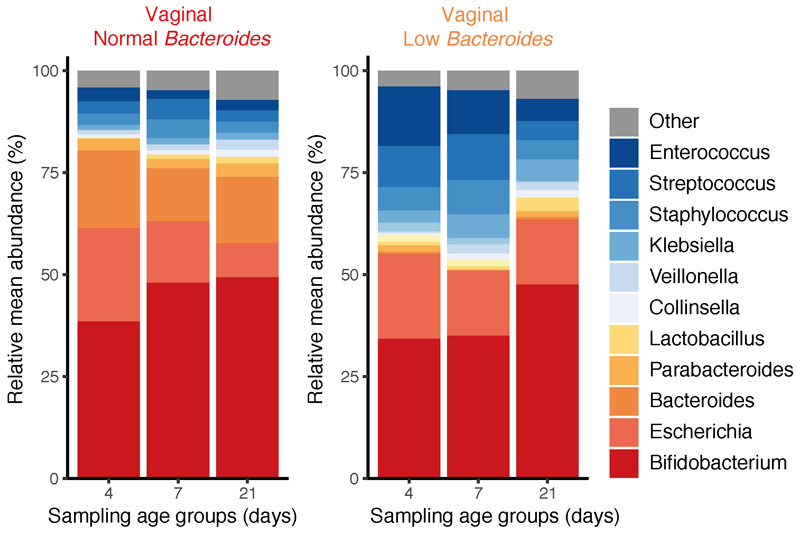
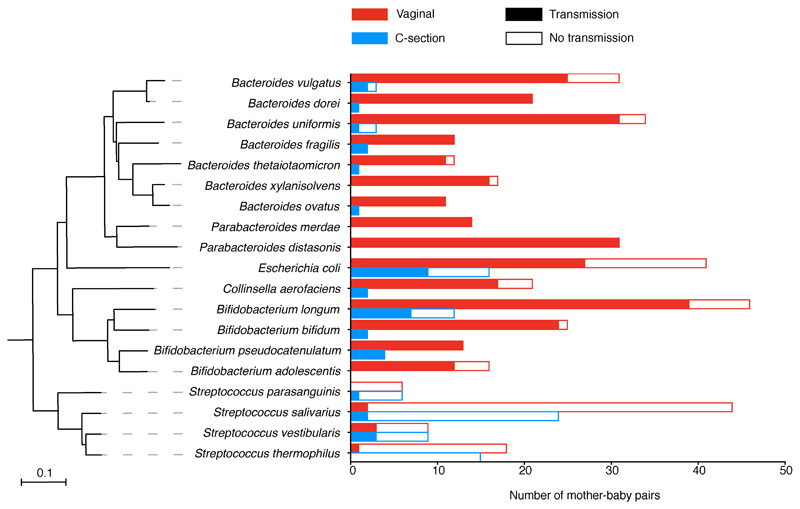
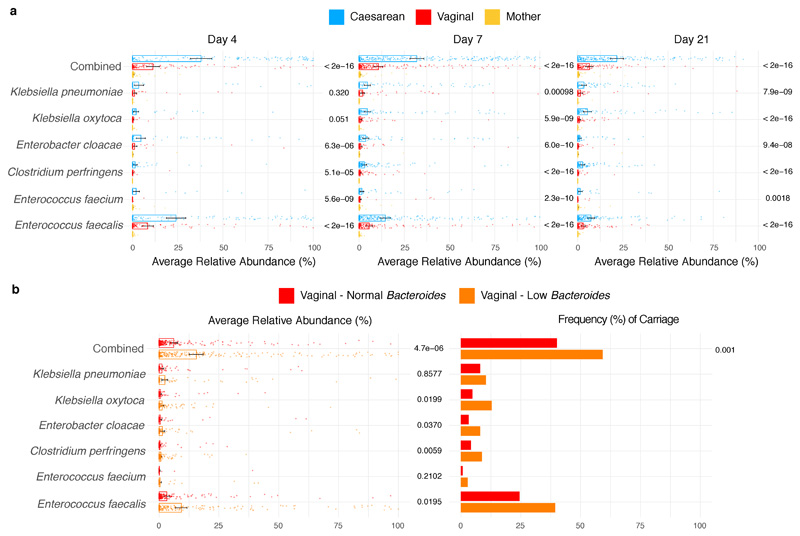
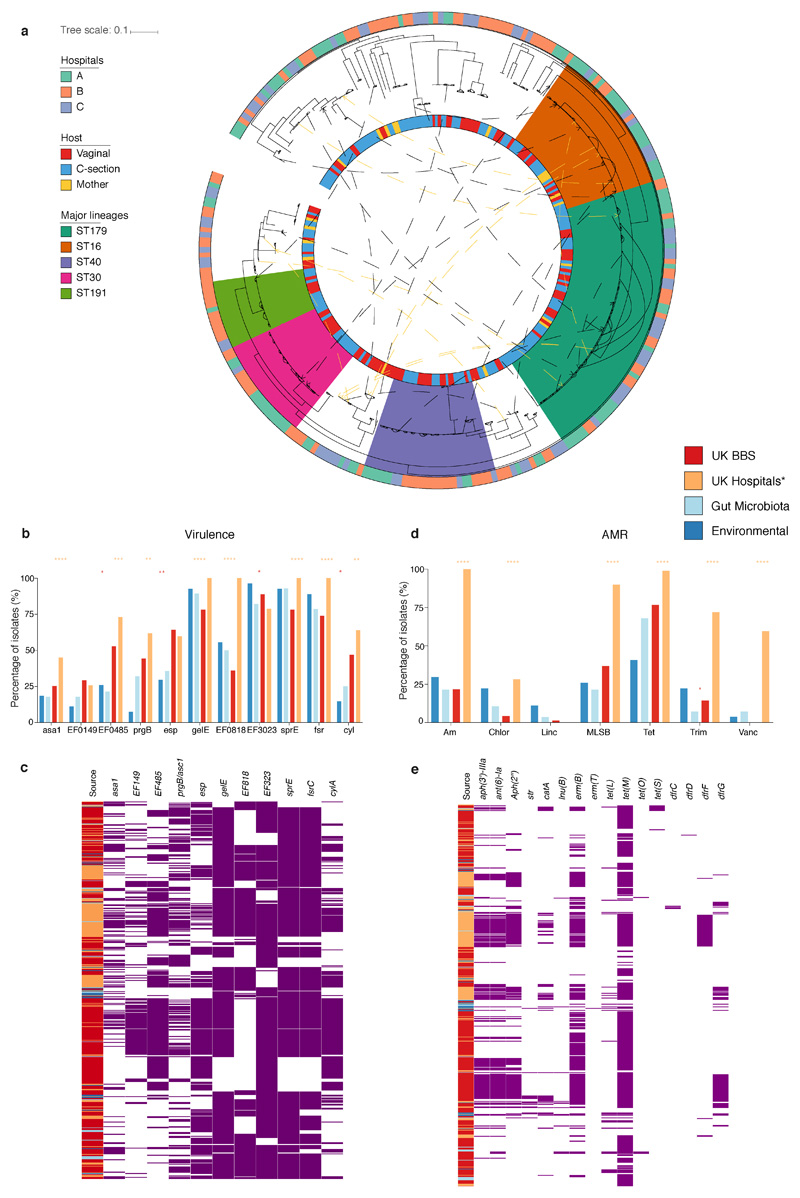
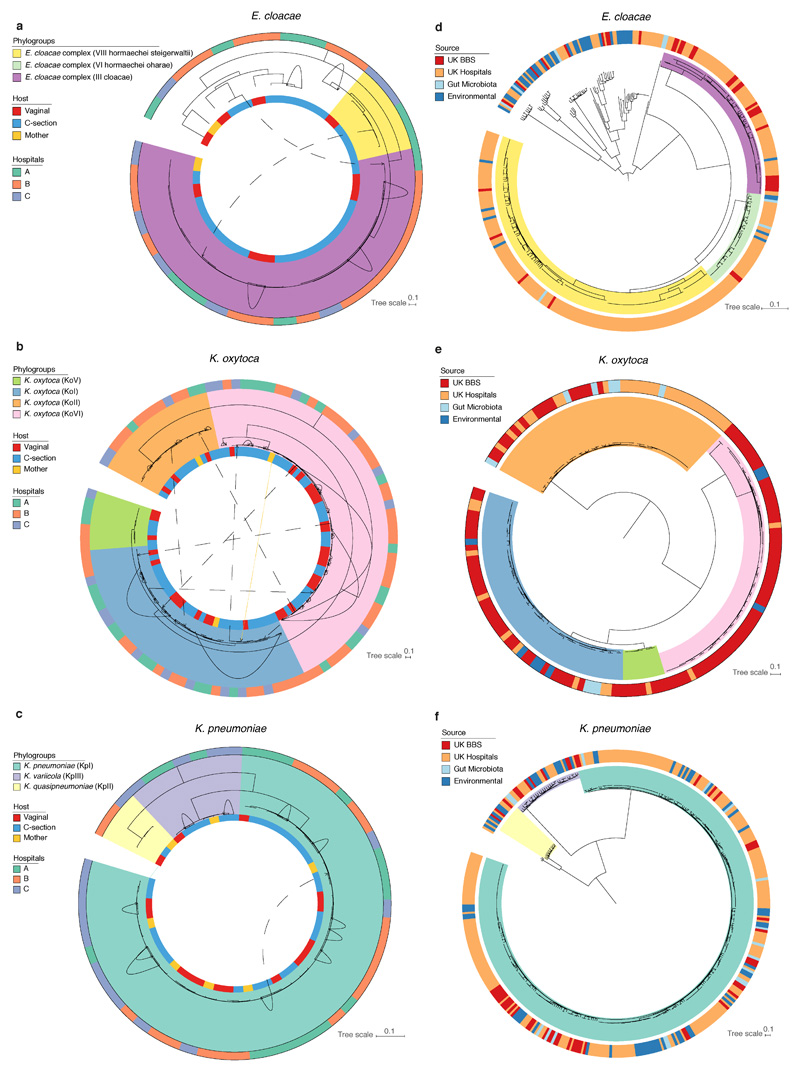
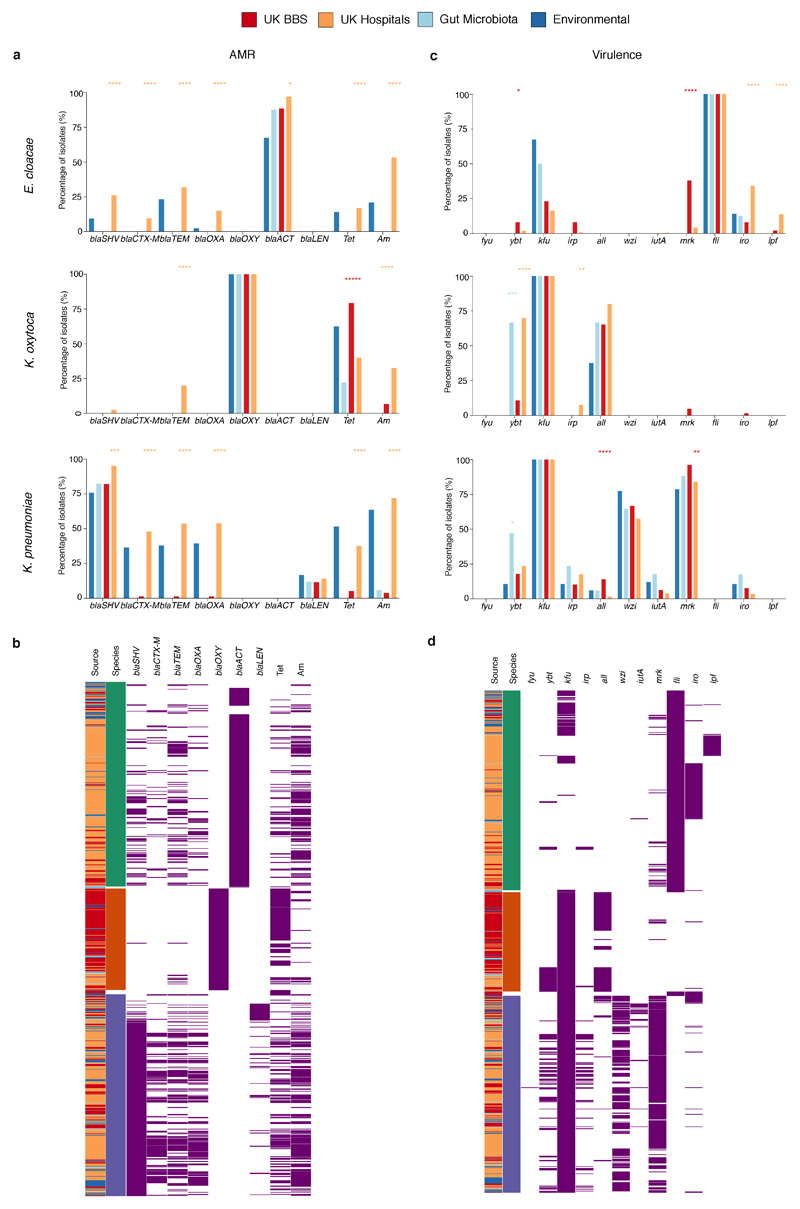
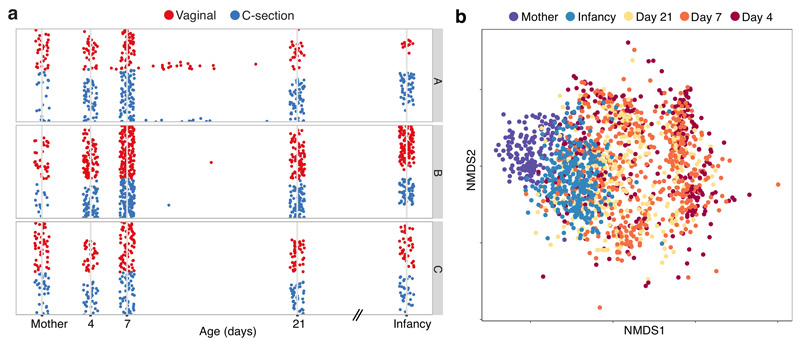
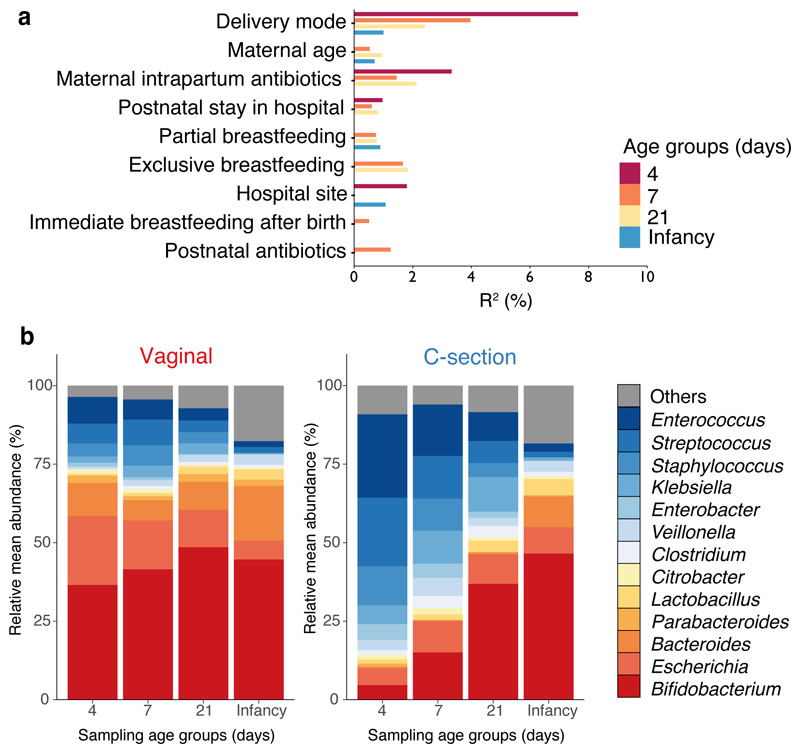
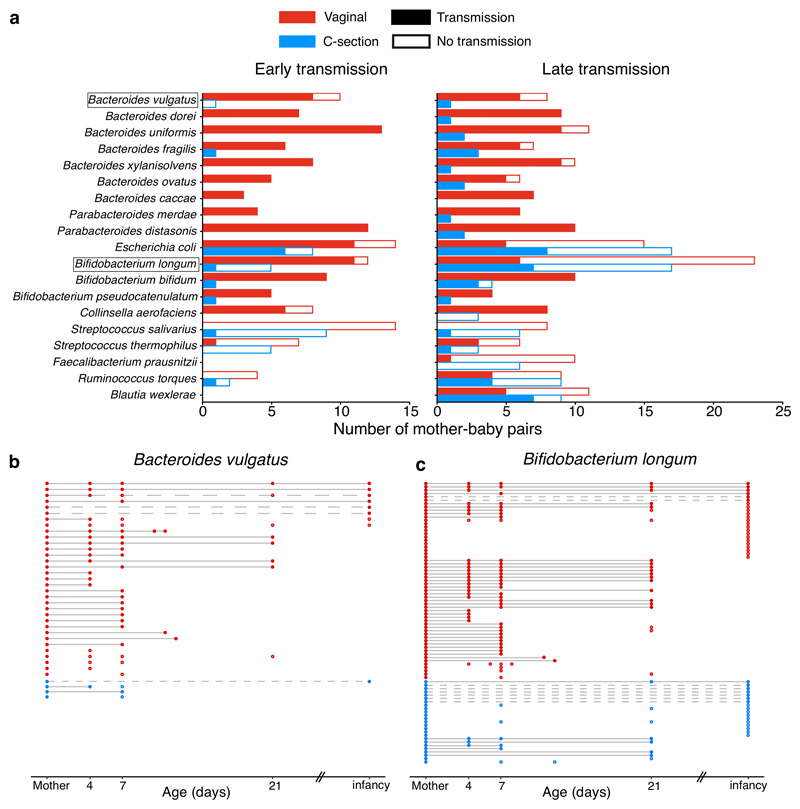
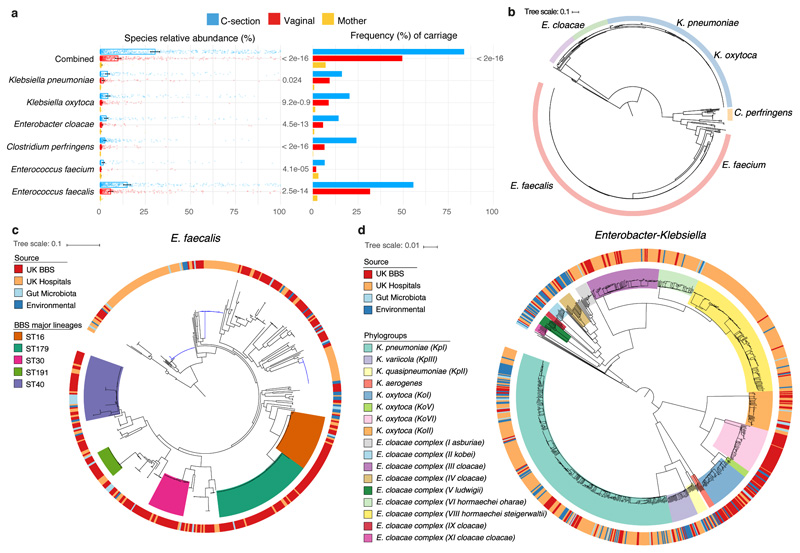
Immediately after birth, newborn babies experience rapid colonization by microorganisms from their mothers and the surrounding environment1. Diseases in childhood and later in life are potentially mediated by the perturbation of the colonization of the infant gut microbiota2. However, the effects of delivery via caesarean section on the earliest stages of the acquisition and development of the gut microbiota, during the neonatal period (≤1 month), remain controversial3,4. Here we report the disrupted transmission of maternal Bacteroides strains, and high-level colonization by opportunistic pathogens associated with the hospital environment (including Enterococcus, Enterobacter and Klebsiella species), in babies delivered by caesarean section. These effects were also seen, to a lesser extent, in vaginally delivered babies whose mothers underwent antibiotic prophylaxis and in babies who were not breastfed during the neonatal period. We applied longitudinal sampling and whole-genome shotgun metagenomic analysis to 1,679 gut microbiota samples (taken at several time points during the neonatal period, and in infancy) from 596 full-term babies born in UK hospitals; for a subset of these babies, we collected additional matched samples from mothers (175 mothers paired with 178 babies). This analysis demonstrates that the mode of delivery is a significant factor that affects the composition of the gut microbiota throughout the neonatal period, and into infancy. Matched large-scale culturing and whole-genome sequencing of over 800 bacterial strains from these babies identified virulence factors and clinically relevant antimicrobial resistance in opportunistic pathogens that may predispose individuals to opportunistic infections. Our findings highlight the critical role of the local environment in establishing the gut microbiota in very early life, and identify colonization with antimicrobial-resistance-containing opportunistic pathogens as a previously underappreciated risk factor in hospital births.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Tamburini S, Shen N, Wu HC, Clemente JC. The microbiome in early life: implications for health outcomes. Nat Med. 2016;22:713–722. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
