

Polyploidy in creosote bush (Larrea tridentata) shapes the biogeography of specialist herbivores
- PMID: 31534296
- PMCID: PMC6749999
- DOI: 10.1111/jbi.13490
Polyploidy in creosote bush (Larrea tridentata) shapes the biogeography of specialist herbivores
Abstract
Aim: Whole-genome duplication (polyploidy) can influence the biogeography and ecology of plants that differ in ploidy level (cytotype). Here, we address how two consequences of plant polyploidy (parapatry of cytotypes and altered species interactions) shape the biogeography of herbivorous insects.
Location: Warm deserts of North America.
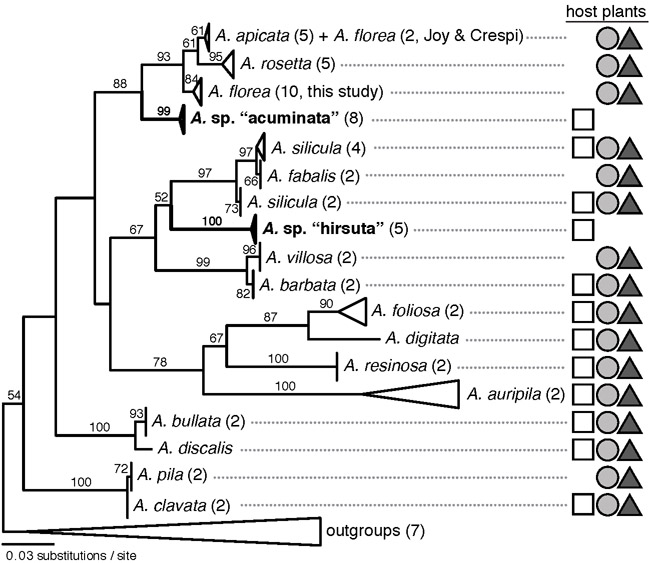
Taxa: Gall midges (Asphondylia auripila group, Diptera: Cecidomyiidae) that attack three parapatric cytotypes of creosote bush (Larrea tridentata, Zygophyllaceae).
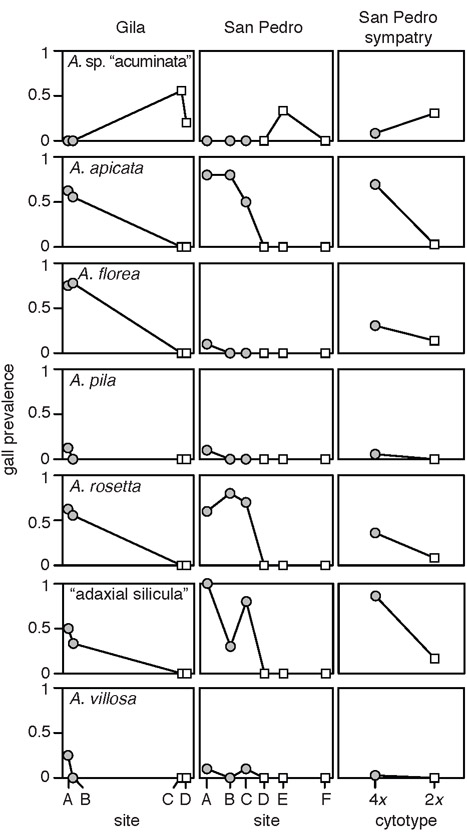
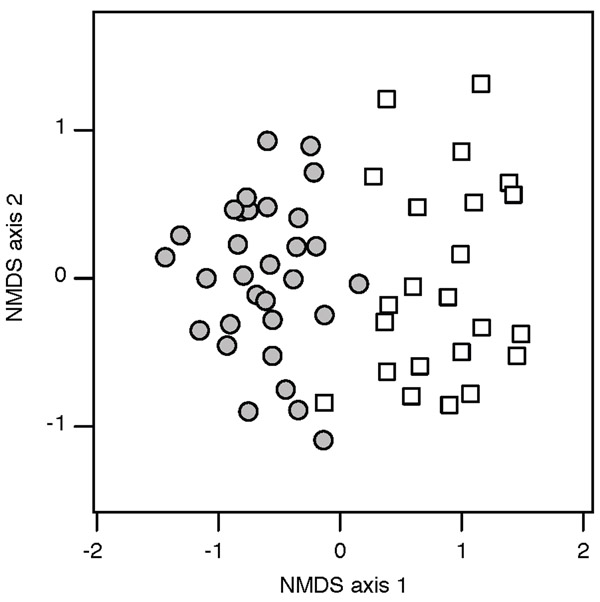
Methods: We surveyed Asphondylia species diversity at 177 sites across a 2300-km extent. After noting a correspondence between the distributions of eight Asphondylia species and L. tridentata cytotypes, we fine-mapped Asphondylia species range limits with transects spanning cytotype contact zones. We then tested whether plant-insect interactions and/or abiotic factors explain this coincidence by (1) comparing attack rates and gall midge communities on alternative cytotypes in a narrow zone of sympatry and (2) using species distribution models (SDMs) to determine if climatically suitable habitat for each midge species extended beyond cytotype contact zones.
Results: The range limits of 6/17 Asphondylia species (including two novel putative species confirmed with COI sequencing) perfectly coincided with the contact zone of diploid and tetraploid L. tridentata. One midge species was restricted to diploid host plants while five were restricted to tetraploid and hexaploid host plants. Where diploid and tetraploid L. tridentata are sympatric, cytotype-restricted midge species more frequently attacked their typical host and Asphondylia community structure differed markedly between cytotypes. SDMs predicted that distributions of cytotype-restricted midge species were not constrained by climatic conditions near cytotype contact zones.
Main conclusions: Contact zones between plant cytotypes are dispersal barriers for many Asphondylia species due to plant-insect interactions. The distribution of L. tridentata cytotypes therefore shapes herbivore species ranges and herbivore community structure across North American deserts. Our results demonstrate that polyploidy in plants can affect the biogeography of ecological communities.
Keywords: Asphondylia; Cecidomyiidae; Larrea tridentata; North American deserts; creosote bush; gall midge; herbivory; polyploidy; species distribution models; species interactions.
Figures


References
-
- Aiello-Lammens ME, Boria RA, Radosavljevic A, Vilela B, & Anderson RP (2015) spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography, 38, 541–545.
-
- Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecology, 26, 32–46.
-
- Anderson RP, Peterson AT, & Gómez-Laverde M (2002) Using niche-based GIS modeling to test geographic predictions of competitive exclusion and competitive release in South American pocket mice. Oikos, 98, 3–16.
-
- Arvanitis L, Wiklund C, & Ehrlén J (2007) Butterfly seed predation: effects of landscape characteristics, plant ploidy level and population structure. Oecologia, 152, 275–285. - PubMed
-
- Arvanitis L, Wiklund C, Münzbergova Z, Dahlgren JP, & Ehrlén J (2010) Novel antagonistic interactions associated with plant polyploidization influence trait selection and habitat preference. Ecology Letters, 13, 330–337. - PubMed