

Mitochondrial PE potentiates respiratory enzymes to amplify skeletal muscle aerobic capacity
- PMID: 31535029
- PMCID: PMC6739096
- DOI: 10.1126/sciadv.aax8352
Mitochondrial PE potentiates respiratory enzymes to amplify skeletal muscle aerobic capacity
Abstract
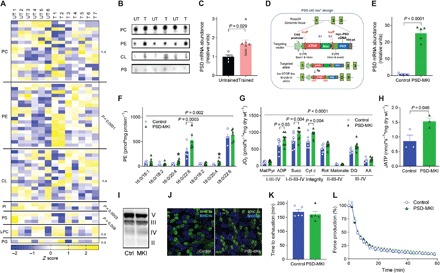
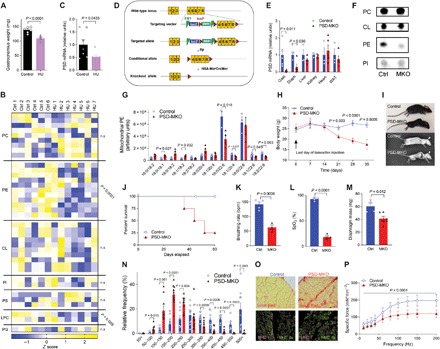
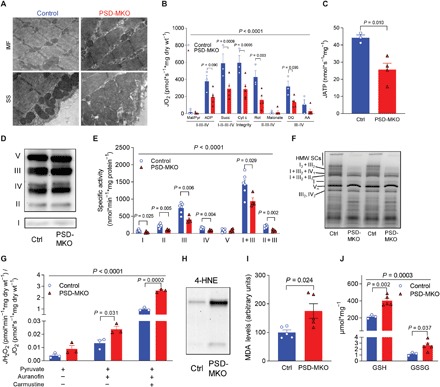
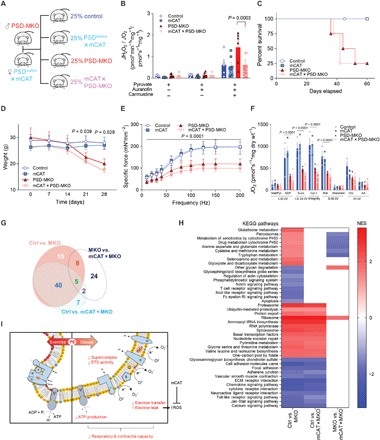
Exercise capacity is a strong predictor of all-cause mortality. Skeletal muscle mitochondrial respiratory capacity, its biggest contributor, adapts robustly to changes in energy demands induced by contractile activity. While transcriptional regulation of mitochondrial enzymes has been extensively studied, there is limited information on how mitochondrial membrane lipids are regulated. Here, we show that exercise training or muscle disuse alters mitochondrial membrane phospholipids including phosphatidylethanolamine (PE). Addition of PE promoted, whereas removal of PE diminished, mitochondrial respiratory capacity. Unexpectedly, skeletal muscle-specific inhibition of mitochondria-autonomous synthesis of PE caused respiratory failure because of metabolic insults in the diaphragm muscle. While mitochondrial PE deficiency coincided with increased oxidative stress, neutralization of the latter did not rescue lethality. These findings highlight the previously underappreciated role of mitochondrial membrane phospholipids in dynamically controlling skeletal muscle energetics and function.
Figures
References
-
- Myers J., Prakash M., Froelicher V., Do D., Partington S., Atwood J. E., Exercise capacity and mortality among men referred for exercise testing. N. Engl. J. Med. 346, 793–801 (2002). - PubMed
-
- Bassett D. R. Jr., Howley E. T., Limiting factors for maximum oxygen uptake and determinants of endurance performance. Med. Sci. Sports Exerc. 32, 70–84 (2000). - PubMed
-
- Holloszy J. O., Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J. Biol. Chem. 242, 2278–2282 (1967). - PubMed
-
- Henriksson J., Reitman J. S., Time course of changes in human skeletal muscle succinate dehydrogenase and cytochrome oxidase activities and maximal oxygen uptake with physical activity and inactivity. Acta Physiol. Scand. 99, 91–97 (1977). - PubMed
-
- Lin J., Wu H., Tarr P. T., Zhang C.-Y., Wu Z., Boss O., Michael L. F., Puigserver P., Isotani E., Olson E. N., Lowell B. B., Bassel-Duby R., Spieglelman B. M., Transcriptional co-activator PGC-1α drives the formation of slow-twitch muscle fibres. Nature 418, 797–801 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- S10 OD016232/OD/NIH HHS/United States
- R35 GM131854/GM/NIGMS NIH HHS/United States
- R03 DK109888/DK/NIDDK NIH HHS/United States
- P30 DK089503/DK/NIDDK NIH HHS/United States
- P30 AG028740/AG/NIA NIH HHS/United States
- R21 AG063077/AG/NIA NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- R01 DK099034/DK/NIDDK NIH HHS/United States
- U24 DK097153/DK/NIDDK NIH HHS/United States
- R01 AT008375/AT/NCCIH NIH HHS/United States
- L30 DK110338/DK/NIDDK NIH HHS/United States
- F32 DK109556/DK/NIDDK NIH HHS/United States
- R01 DK110656/DK/NIDDK NIH HHS/United States
- R01 HL123647/HL/NHLBI NIH HHS/United States
- R01 DK107397/DK/NIDDK NIH HHS/United States
- R01 AG050781/AG/NIA NIH HHS/United States
- U54 DK110858/DK/NIDDK NIH HHS/United States
- R01 AR071967/AR/NIAMS NIH HHS/United States
- P30 DK020579/DK/NIDDK NIH HHS/United States
- P30 DK056350/DK/NIDDK NIH HHS/United States
- F32 HL129632/HL/NHLBI NIH HHS/United States
- S10 OD021505/OD/NIH HHS/United States
- K01 DK095774/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
