

A distinct transition from cell growth to physiological homeostasis in the tendon
- PMID: 31535975
- PMCID: PMC6791717
- DOI: 10.7554/eLife.48689
A distinct transition from cell growth to physiological homeostasis in the tendon
Abstract
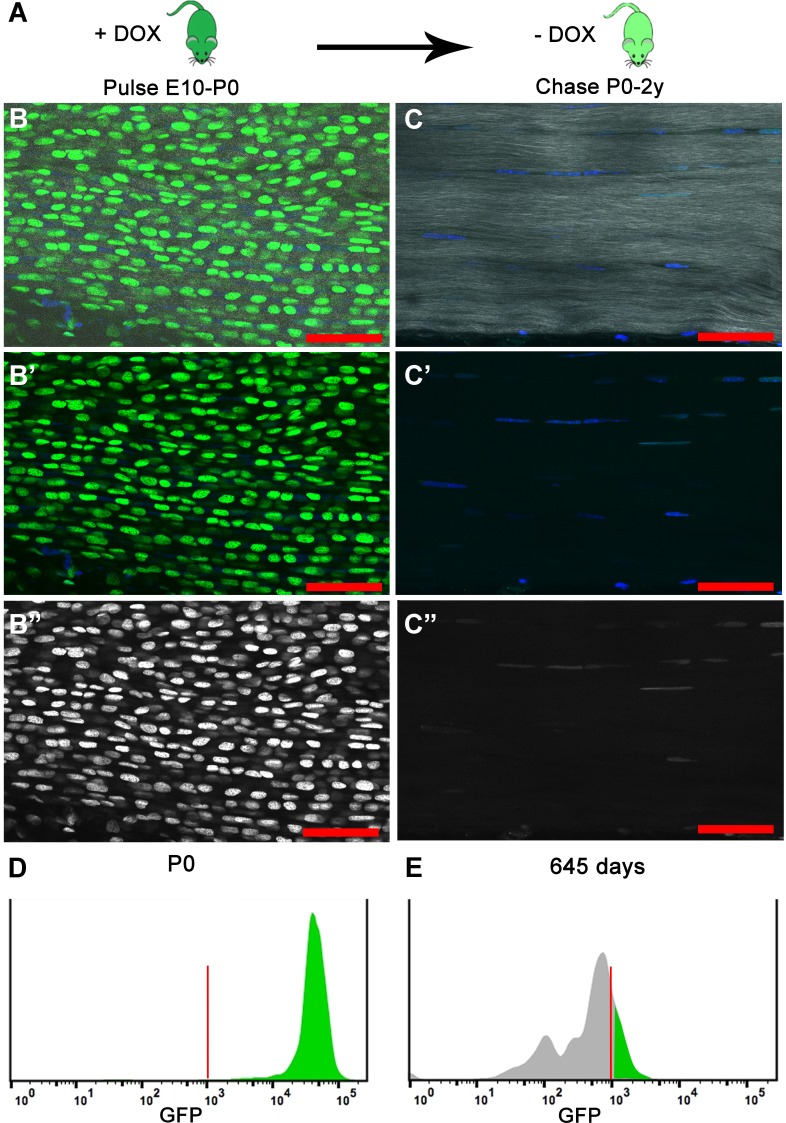
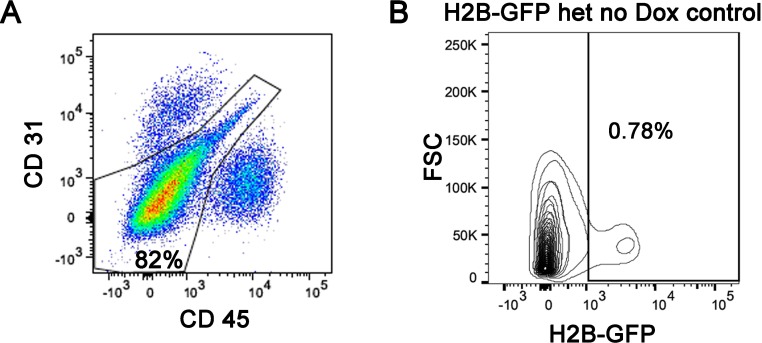
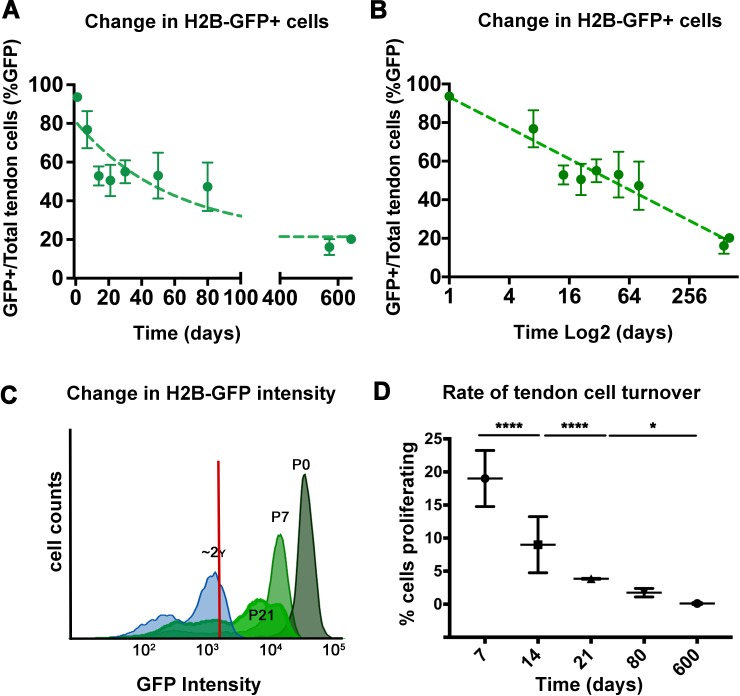
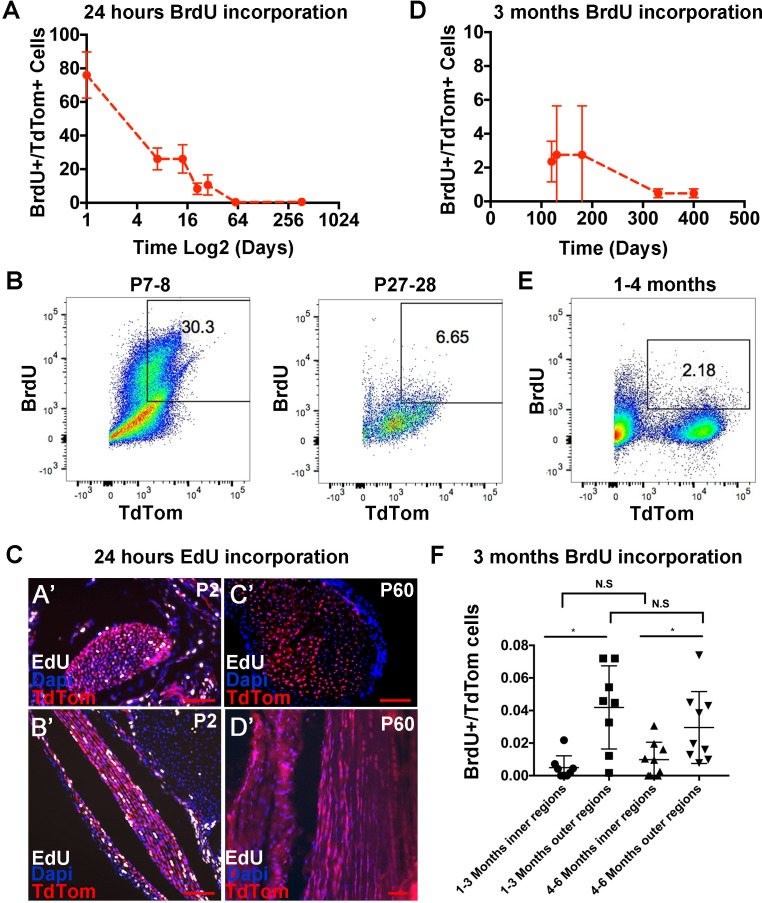
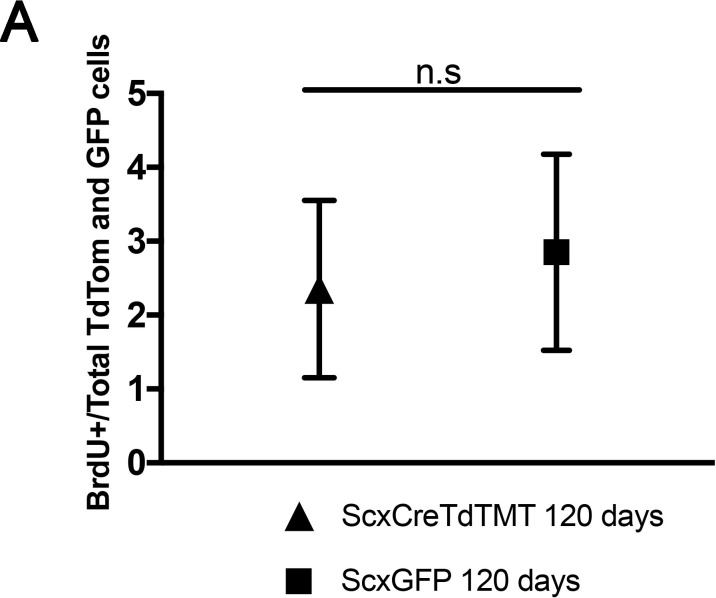
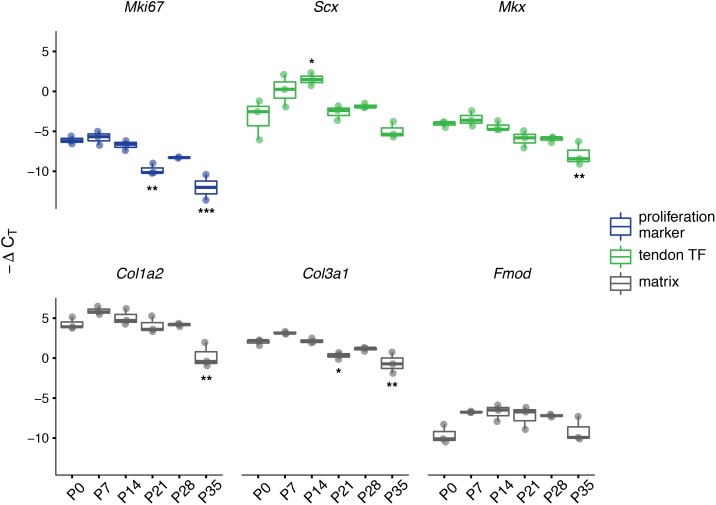
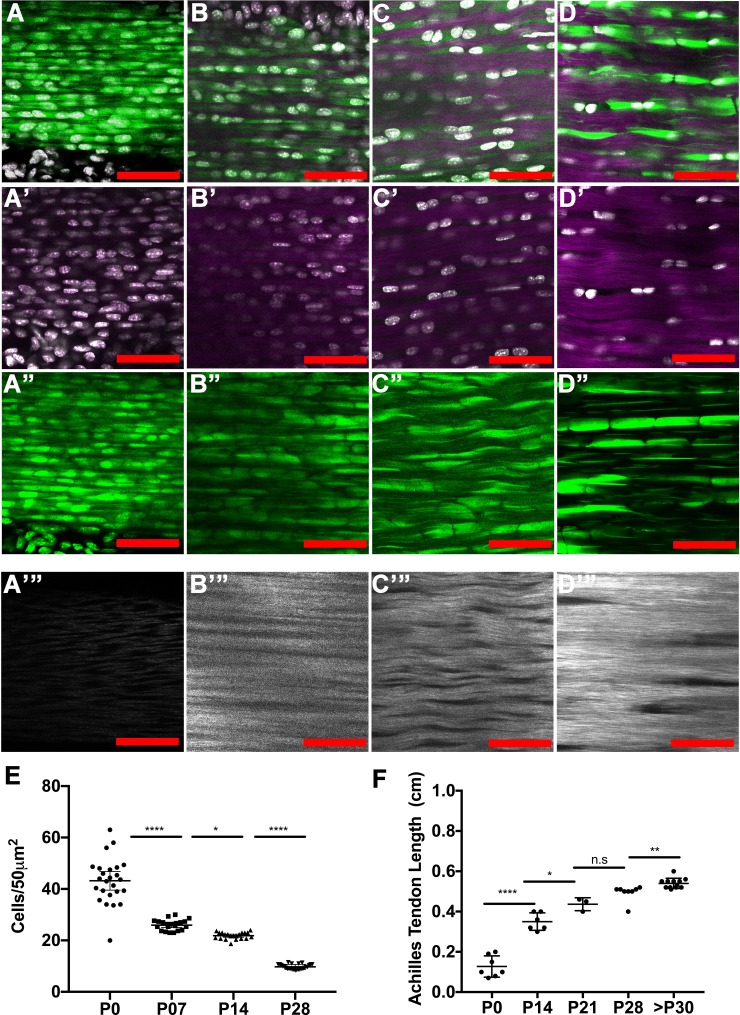
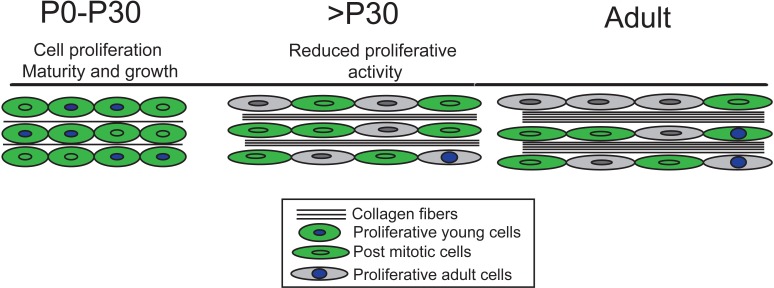
Changes in cell proliferation define transitions from tissue growth to physiological homeostasis. In tendons, a highly organized extracellular matrix undergoes significant postnatal expansion to drive growth, but once formed, it appears to undergo little turnover. However, tendon cell activity during growth and homeostatic maintenance is less well defined. Using complementary methods of genetic H2B-GFP pulse-chase labeling and BrdU incorporation in mice, we show significant postnatal tendon cell proliferation, correlating with longitudinal Achilles tendon growth. Around day 21, there is a transition in cell turnover with a significant decline in proliferation. After this time, we find low amounts of homeostatic tendon cell proliferation from 3 to 20 months. These results demonstrate that tendons harbor significant postnatal mitotic activity, and limited, but detectable activity in adult and aged stages. It also points towards the possibility that the adult tendon harbors resident tendon progenitor populations, which would have important therapeutic implications.
Keywords: developmental biology; homeostasis; mouse; musculoskeletal; regenerative medicine; stem cells; tendon; tissue growth.
© 2019, Grinstein et al.
Conflict of interest statement
MG, HD, LO, KZ, TC, JG No competing interests declared
Figures
References
-
- Banes AJ, Donlon K, Link GW, Gillespie Y, Bevin AG, Peterson HD, Bynum D, Watts S, Dahners L. Cell populations of tendon: a simplified method for isolation of synovial cells and internal fibroblasts: confirmation of origin and biologic properties. Journal of Orthopaedic Research. 1988;6:83–94. doi: 10.1002/jor.1100060111. - DOI - PubMed
-
- Bassat E, Mutlak YE, Genzelinakh A, Shadrin IY, Baruch Umansky K, Yifa O, Kain D, Rajchman D, Leach J, Riabov Bassat D, Udi Y, Sarig R, Sagi I, Martin JF, Bursac N, Cohen S, Tzahor E. The extracellular matrix protein agrin promotes heart regeneration in mice. Nature. 2017;547:179–184. doi: 10.1038/nature22978. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
