

PD-1hiCXCR5- T peripheral helper cells promote B cell responses in lupus via MAF and IL-21
- PMID: 31536480
- PMCID: PMC6824311
- DOI: 10.1172/jci.insight.130062
PD-1hiCXCR5- T peripheral helper cells promote B cell responses in lupus via MAF and IL-21
Abstract
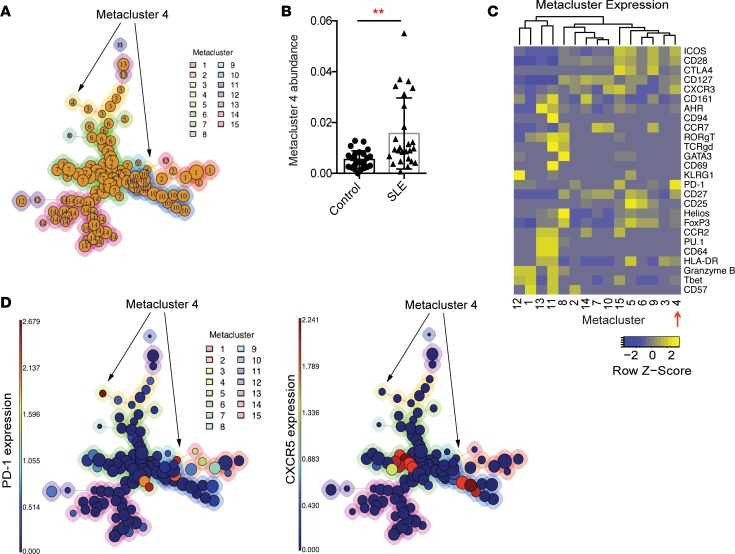
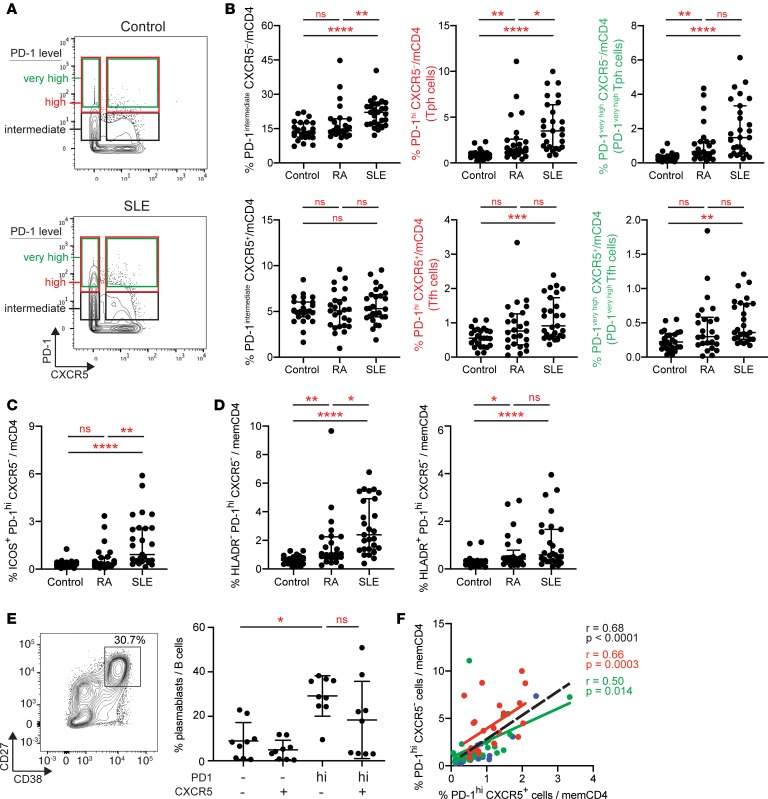
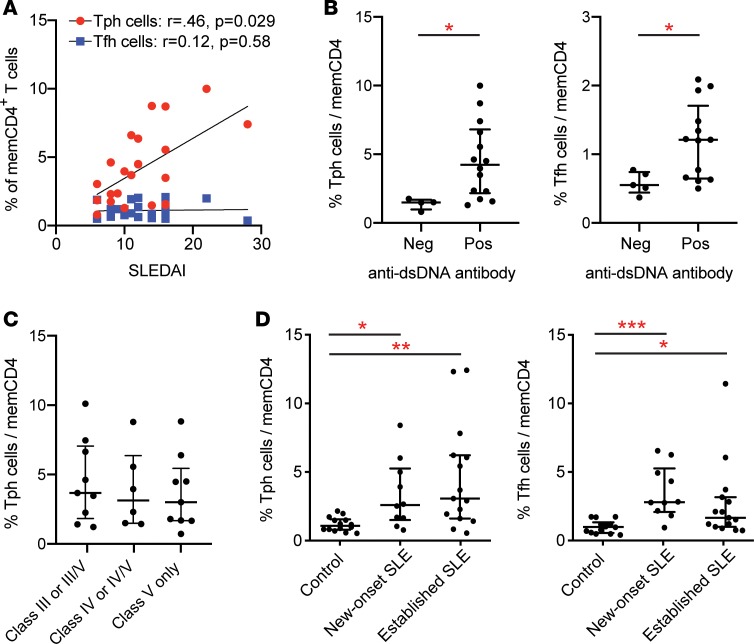
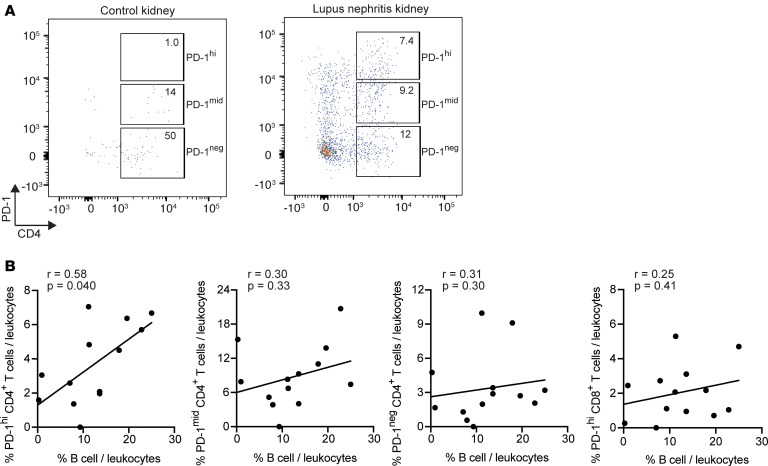
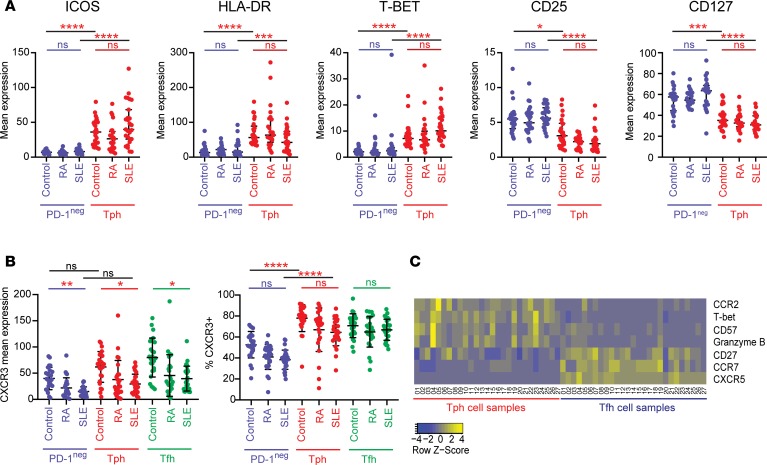
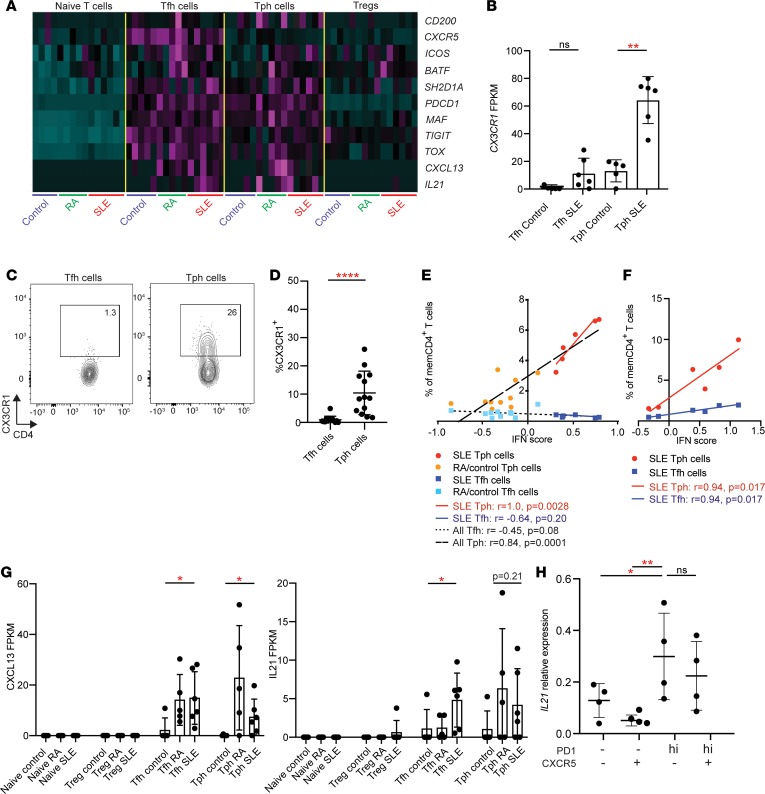
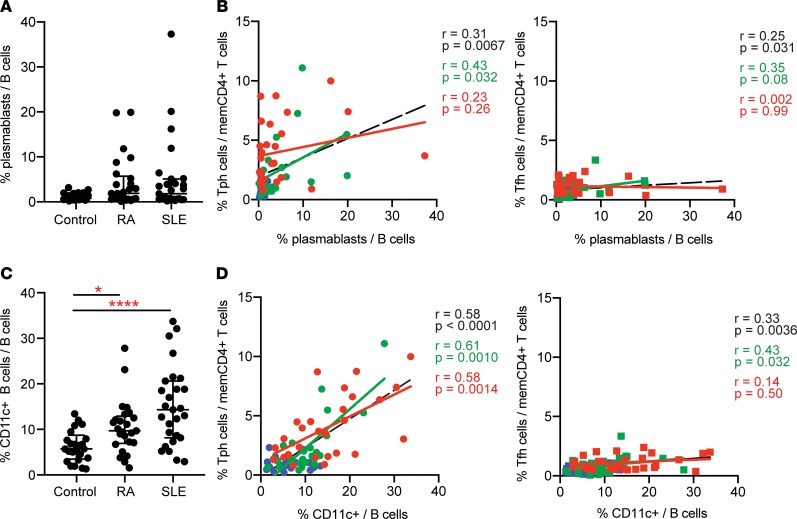
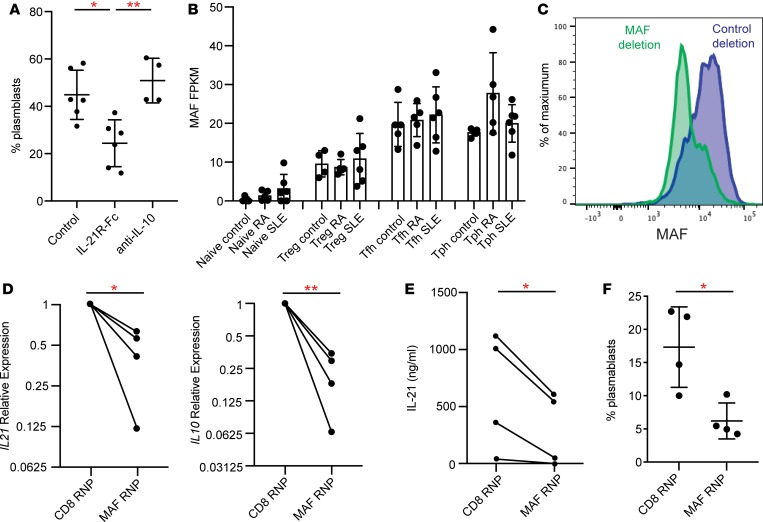
Systemic lupus erythematosus (SLE) is an autoimmune disease characterized by pathologic T cell-B cell interactions and autoantibody production. Defining the T cell populations that drive B cell responses in SLE may enable design of therapies that specifically target pathologic cell subsets. Here, we evaluated the phenotypes of CD4+ T cells in the circulation of 52 SLE patients drawn from multiple cohorts and identified a highly expanded PD-1hiCXCR5-CD4+ T cell population. Cytometric, transcriptomic, and functional assays demonstrated that PD-1hiCXCR5-CD4+ T cells from SLE patients are T peripheral helper (Tph) cells, a CXCR5- T cell population that stimulates B cell responses via IL-21. The frequency of Tph cells, but not T follicular helper (Tfh) cells, correlated with both clinical disease activity and the frequency of CD11c+ B cells in SLE patients. PD-1hiCD4+ T cells were found within lupus nephritis kidneys and correlated with B cell numbers in the kidney. Both IL-21 neutralization and CRISPR-mediated deletion of MAF abrogated the ability of Tph cells to induce memory B cell differentiation into plasmablasts in vitro. These findings identify Tph cells as a highly expanded T cell population in SLE and suggest a key role for Tph cells in stimulating pathologic B cell responses.
Keywords: Adaptive immunity; Autoimmunity; Immunology; Lupus; T cells.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
