

MMP9 inhibition increases erythropoiesis in RPS14-deficient del(5q) MDS models through suppression of TGF-β pathways
- PMID: 31540902
- PMCID: PMC6759738
- DOI: 10.1182/bloodadvances.2019000537
MMP9 inhibition increases erythropoiesis in RPS14-deficient del(5q) MDS models through suppression of TGF-β pathways
Abstract
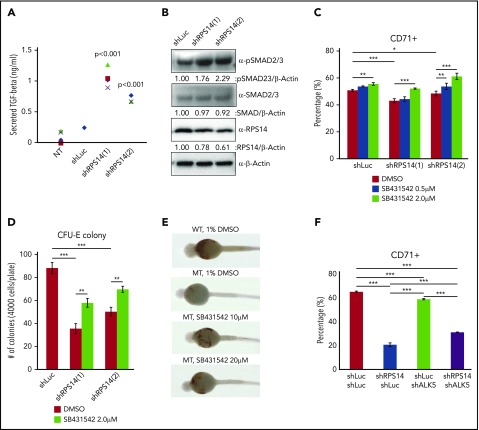
The del(5q) myelodysplastic syndrome (MDS) is a distinct subtype of MDS, associated with deletion of the ribosomal protein S14 (RPS14) gene that results in macrocytic anemia. This study sought to identify novel targets for the treatment of patients with del(5q) MDS by performing an in vivo drug screen using an rps14-deficient zebrafish model. From this, we identified the secreted gelatinase matrix metalloproteinase 9 (MMP9). MMP9 inhibitors significantly improved the erythroid defect in rps14-deficient zebrafish. Similarly, treatment with MMP9 inhibitors increased the number of colony forming unit-erythroid colonies and the CD71+ erythroid population from RPS14 knockdown human BMCD34+ cells. Importantly, we found that MMP9 expression is upregulated in RPS14-deficient cells by monocyte chemoattractant protein 1. Double knockdown of MMP9 and RPS14 increased the CD71+ population compared with RPS14 single knockdown, suggesting that increased expression of MMP9 contributes to the erythroid defect observed in RPS14-deficient cells. In addition, transforming growth factor β (TGF-β) signaling is activated in RPS14 knockdown cells, and treatment with SB431542, a TGF-β inhibitor, improved the defective erythroid development of RPS14-deficient models. We found that recombinant MMP9 treatment decreases the CD71+ population through increased SMAD2/3 phosphorylation, suggesting that MMP9 directly activates TGF-β signaling in RPS14-deficient cells. Finally, we confirmed that MMP9 inhibitors reduce SMAD2/3 phosphorylation in RPS14-deficient cells to rescue the erythroid defect. In summary, these study results support a novel role for MMP9 in the pathogenesis of del(5q) MDS and the potential for the clinical use of MMP9 inhibitors in the treatment of patients with del(5q) MDS.
© 2019 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures





Similar articles
-
Evolution of severe (transfusion-dependent) anaemia in myelodysplastic syndromes with 5q deletion is characterized by a macrophage-associated failure of the eythropoietic niche.Br J Haematol. 2022 Jul;198(1):114-130. doi: 10.1111/bjh.18163. Epub 2022 Apr 1. Br J Haematol. 2022. PMID: 35362549
-
Low RPS14 expression in MDS without 5q - aberration confers higher apoptosis rate of nucleated erythrocytes and predicts prolonged survival and possible response to lenalidomide in lower risk non-5q- patients.Eur J Haematol. 2013 Jun;90(6):486-93. doi: 10.1111/ejh.12105. Epub 2013 Apr 23. Eur J Haematol. 2013. PMID: 23506134 Clinical Trial.
-
Rps14 haploinsufficiency causes a block in erythroid differentiation mediated by S100A8 and S100A9.Nat Med. 2016 Mar;22(3):288-97. doi: 10.1038/nm.4047. Epub 2016 Feb 15. Nat Med. 2016. PMID: 26878232 Free PMC article.
-
PP2A: The Achilles Heal in MDS with 5q Deletion.Front Oncol. 2014 Sep 23;4:264. doi: 10.3389/fonc.2014.00264. eCollection 2014. Front Oncol. 2014. PMID: 25295231 Free PMC article. Review.
-
Dyserythropoiesis of myelodysplastic syndromes.Curr Opin Hematol. 2017 May;24(3):191-197. doi: 10.1097/MOH.0000000000000325. Curr Opin Hematol. 2017. PMID: 28072603 Review.
Cited by
-
Single-cell transcriptional profile of CD34+ hematopoietic progenitor cells from del(5q) myelodysplastic syndromes and impact of lenalidomide.Nat Commun. 2024 Jun 20;15(1):5272. doi: 10.1038/s41467-024-49529-x. Nat Commun. 2024. PMID: 38902243 Free PMC article.
-
Clinical characteristics of thyroid eye disease and expression profile of peripheral blood immune cells.Sci Rep. 2025 Aug 6;15(1):28666. doi: 10.1038/s41598-025-08904-4. Sci Rep. 2025. PMID: 40764715 Free PMC article.
-
Nucleolar Protein 56 Deficiency in Zebrafish Leads to Developmental Abnormalities and Anemia via p53 and JAK2-STAT3 Signaling.Biology (Basel). 2023 Mar 31;12(4):538. doi: 10.3390/biology12040538. Biology (Basel). 2023. PMID: 37106740 Free PMC article.
-
Sex Effect on Cardiac Damage in Mice With Experimental Autoimmune Encephalomyelitis.ASN Neuro. 2021 Jan-Dec;13:1759091421991771. doi: 10.1177/1759091421991771. ASN Neuro. 2021. PMID: 33541127 Free PMC article.
-
The Dynamic Interface Between the Bone Marrow Vascular Niche and Hematopoietic Stem Cells in Myeloid Malignancy.Front Cell Dev Biol. 2021 Mar 11;9:635189. doi: 10.3389/fcell.2021.635189. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33777944 Free PMC article. Review.
References
-
- Boultwood J, Pellagatti A, McKenzie AN, Wainscoat JS. Advances in the 5q- syndrome. Blood. 2010;116(26):5803-5811. - PubMed
-
- Van den Berghe H, Cassiman JJ, David G, et al. . Distinct haematological disorder with deletion of long arm of no. 5 chromosome. Nature. 1974;251(5474):437-438. - PubMed
-
- Giagounidis AA, Germing U, Haase S, et al. . Clinical, morphological, cytogenetic, and prognostic features of patients with myelodysplastic syndromes and del(5q) including band q31. Leukemia. 2004;18(1):113-119. - PubMed
-
- Boultwood J, Lewis S, Wainscoat JS. The 5q-syndrome. Blood. 1994;84(10):3253-3260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
