

A fungal pathogen deploys a small silencing RNA that attenuates mosquito immunity and facilitates infection
- PMID: 31541102
- PMCID: PMC6754459
- DOI: 10.1038/s41467-019-12323-1
A fungal pathogen deploys a small silencing RNA that attenuates mosquito immunity and facilitates infection
Abstract
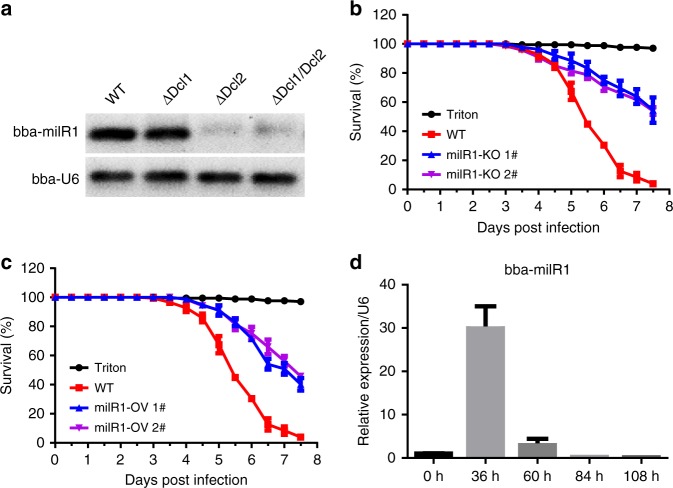
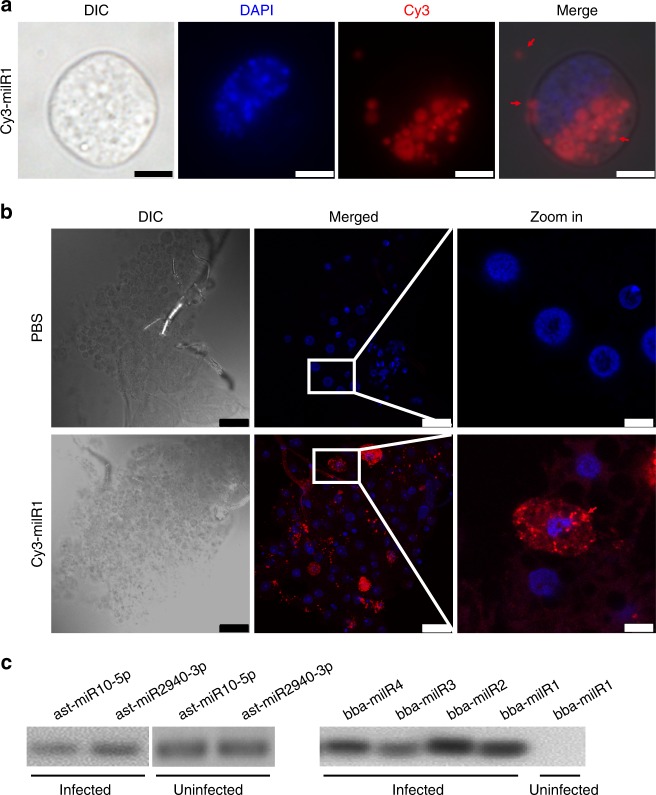
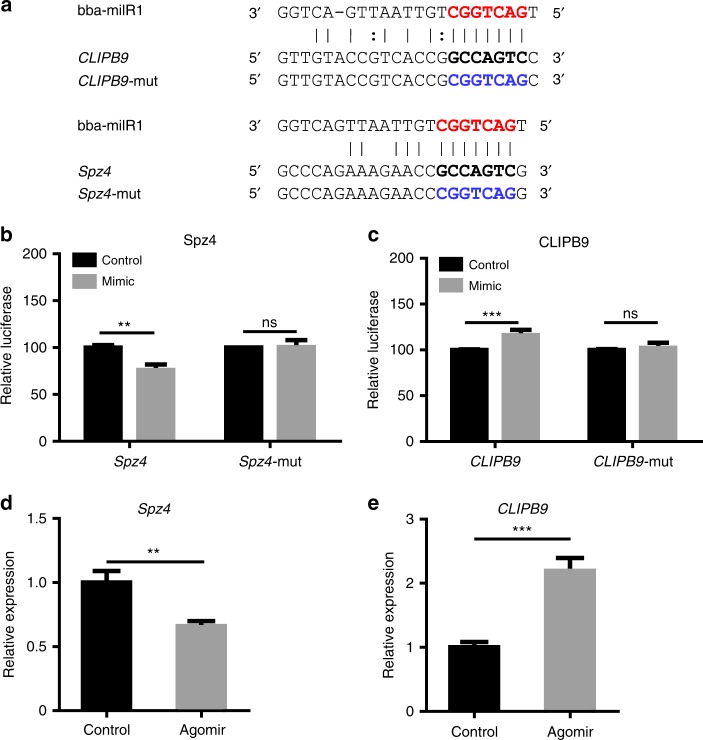
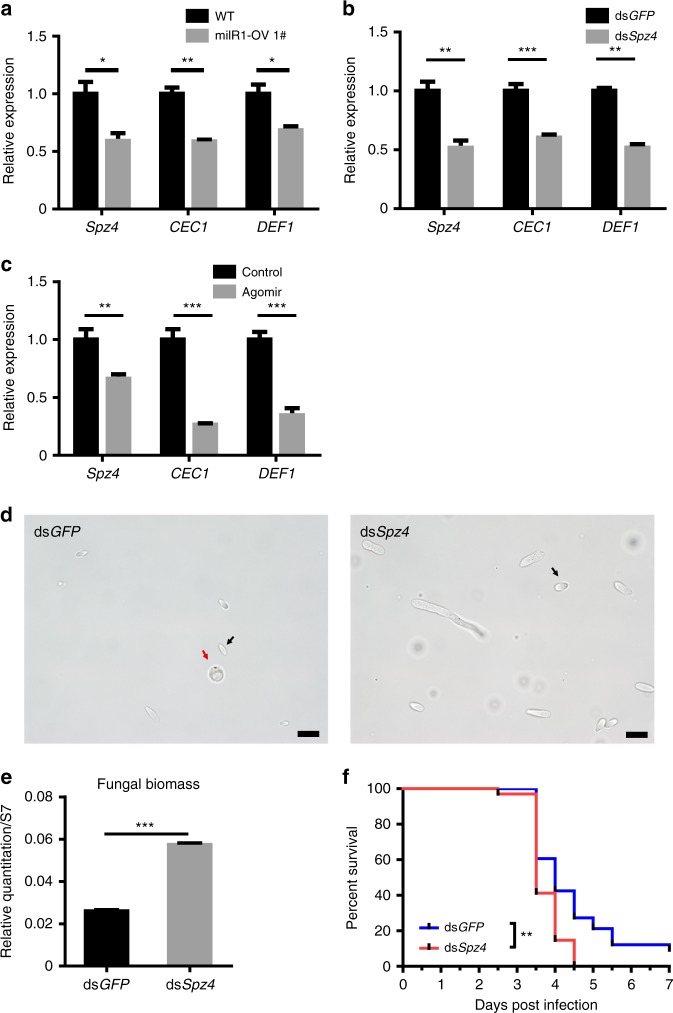
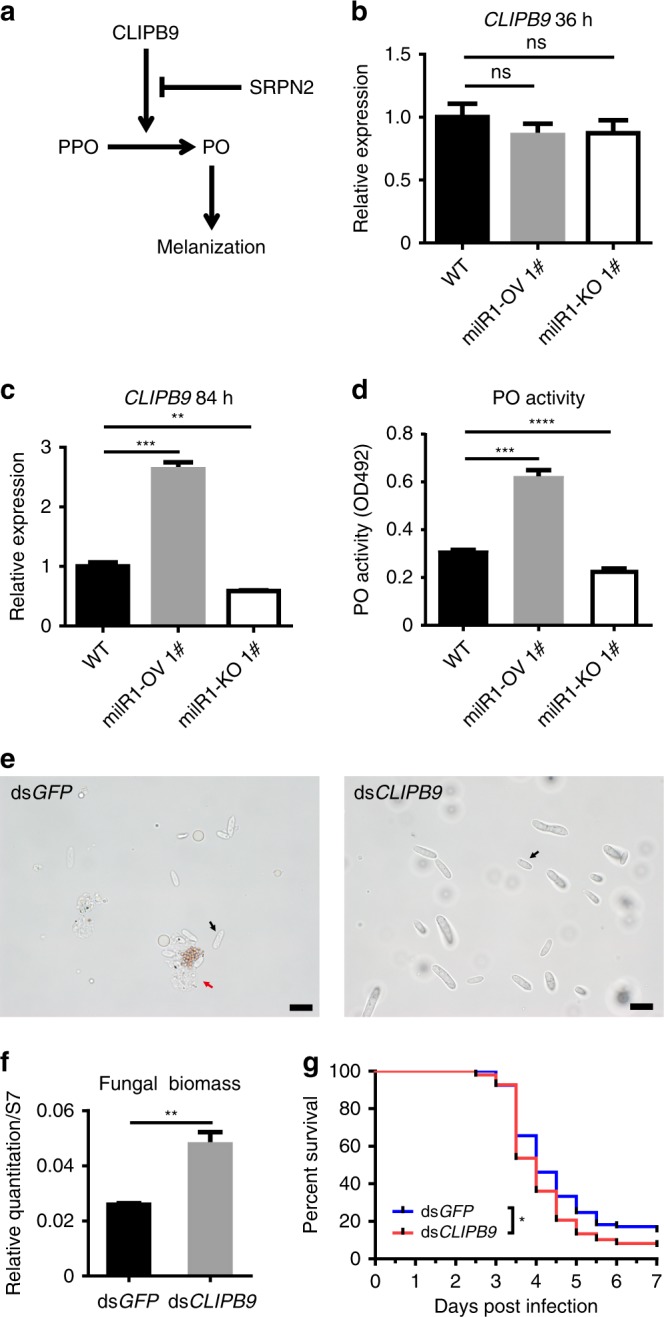
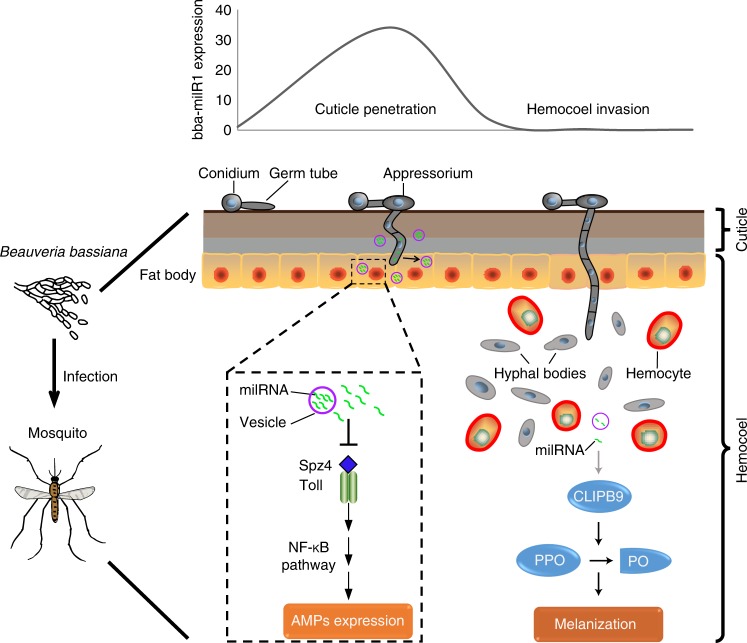
Insecticidal fungi represent a promising alternative to chemical pesticides for disease vector control. Here, we show that the pathogenic fungus Beauveria bassiana exports a microRNA-like RNA (bba-milR1) that hijacks the host RNA-interference machinery in mosquito cells by binding to Argonaute 1 (AGO1). bba-milR1 is highly expressed during fungal penetration of the mosquito integument, and suppresses host immunity by silencing expression of the mosquito Toll receptor ligand Spätzle 4 (Spz4). Later, upon entering the hemocoel, bba-milR1 expression is decreased, which avoids induction of the host proteinase CLIPB9 that activates the melanization response. Thus, our results indicate that the pathogen deploys a cross-kingdom small-RNA effector that attenuates host immunity and facilitates infection.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
The interplay between dose and immune system activation determines fungal infection outcome in the African malaria mosquito, Anopheles gambiae.Dev Comp Immunol. 2018 Aug;85:125-133. doi: 10.1016/j.dci.2018.04.008. Epub 2018 Apr 9. Dev Comp Immunol. 2018. PMID: 29649553 Free PMC article.
-
The Aedes aegypti IMD pathway is a critical component of the mosquito antifungal immune response.Dev Comp Immunol. 2019 Jun;95:1-9. doi: 10.1016/j.dci.2018.12.010. Epub 2018 Dec 22. Dev Comp Immunol. 2019. PMID: 30582948
-
In vivo gene expression profiling of the entomopathogenic fungus Beauveria bassiana elucidates its infection stratagems in Anopheles mosquito.Sci China Life Sci. 2017 Aug;60(8):839-851. doi: 10.1007/s11427-017-9101-3. Epub 2017 Jul 27. Sci China Life Sci. 2017. PMID: 28755300
-
NF-κB-Like Signaling Pathway REL2 in Immune Defenses of the Malaria Vector Anopheles gambiae.Front Cell Infect Microbiol. 2017 Jun 21;7:258. doi: 10.3389/fcimb.2017.00258. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28680852 Free PMC article. Review.
-
Insect Immunity to Entomopathogenic Fungi.Adv Genet. 2016;94:251-85. doi: 10.1016/bs.adgen.2015.11.002. Epub 2016 Jan 22. Adv Genet. 2016. PMID: 27131327 Review.
Cited by
-
Cross-Kingdom Interaction of miRNAs and Gut Microbiota with Non-Invasive Diagnostic and Therapeutic Implications in Colorectal Cancer.Int J Mol Sci. 2023 Jun 23;24(13):10520. doi: 10.3390/ijms241310520. Int J Mol Sci. 2023. PMID: 37445698 Free PMC article. Review.
-
Global discovery, expression pattern, and regulatory role of miRNA-like RNAs in Ascosphaera apis infecting the Asian honeybee larvae.Front Microbiol. 2025 Mar 4;16:1551625. doi: 10.3389/fmicb.2025.1551625. eCollection 2025. Front Microbiol. 2025. PMID: 40104596 Free PMC article.
-
The Aedes aegypti mosquito evolves two types of prophenoloxidases with diversified functions.Proc Natl Acad Sci U S A. 2025 Jan 21;122(3):e2413131122. doi: 10.1073/pnas.2413131122. Epub 2025 Jan 14. Proc Natl Acad Sci U S A. 2025. PMID: 39808654 Free PMC article.
-
Plant mRNAs move into a fungal pathogen via extracellular vesicles to reduce infection.Cell Host Microbe. 2024 Jan 10;32(1):93-105.e6. doi: 10.1016/j.chom.2023.11.020. Epub 2023 Dec 15. Cell Host Microbe. 2024. PMID: 38103543 Free PMC article.
-
Filamentous Fungi Extracellular Vesicles.Curr Top Microbiol Immunol. 2021;432:45-55. doi: 10.1007/978-3-030-83391-6_5. Curr Top Microbiol Immunol. 2021. PMID: 34972877
References
-
- World Health Organization, World Malaria Report (World Health Organization, 2018); www.who.int/malaria/publications/worldmalaria-report-2018/en/.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
