

The internal interaction in RBBP5 regulates assembly and activity of MLL1 methyltransferase complex
- PMID: 31544921
- PMCID: PMC6821195
- DOI: 10.1093/nar/gkz819
The internal interaction in RBBP5 regulates assembly and activity of MLL1 methyltransferase complex
Abstract
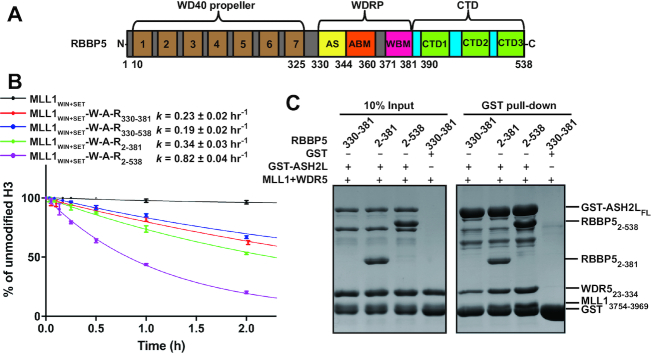
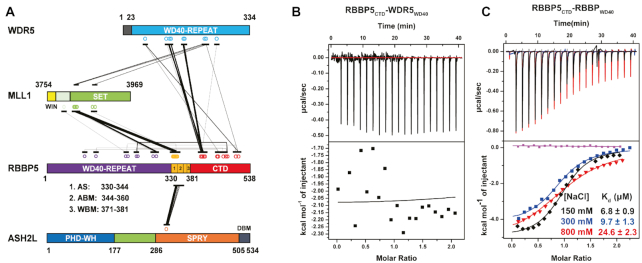
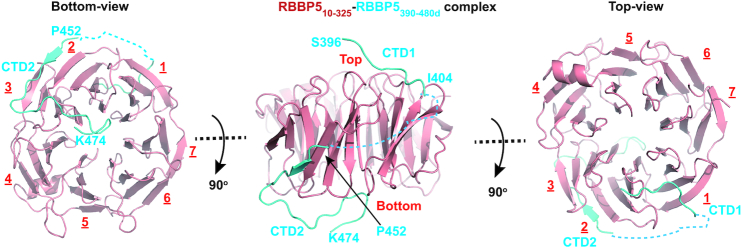
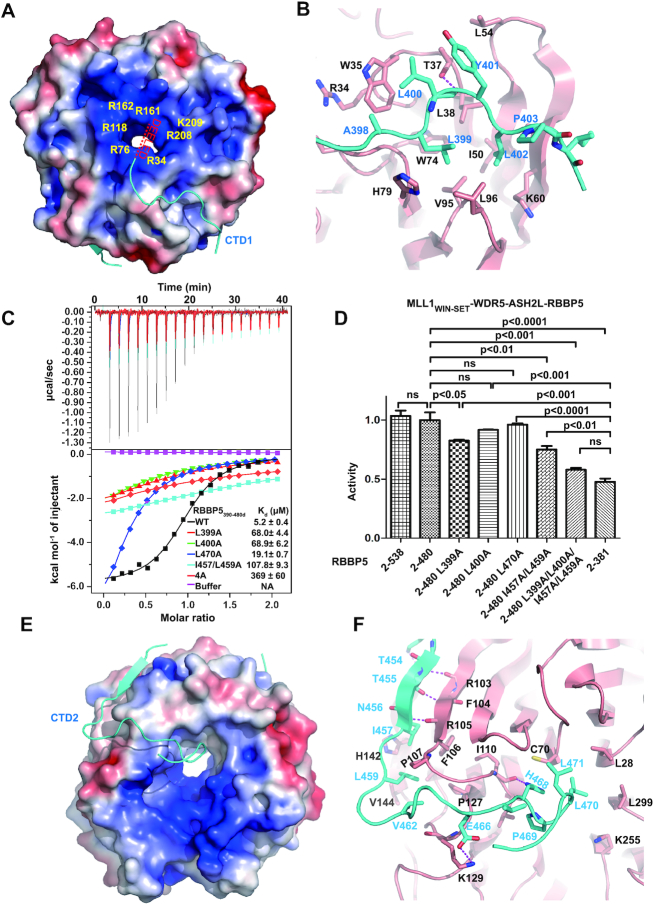
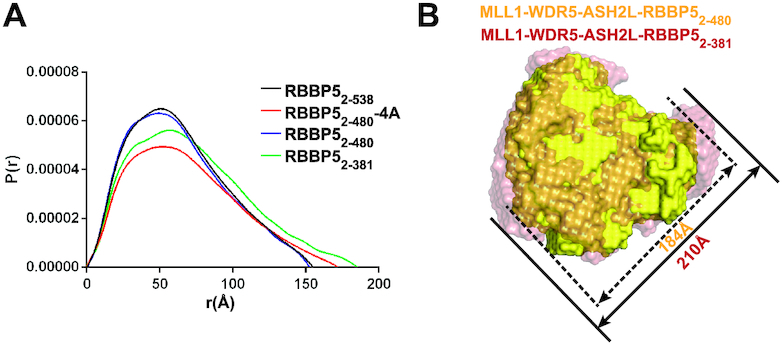
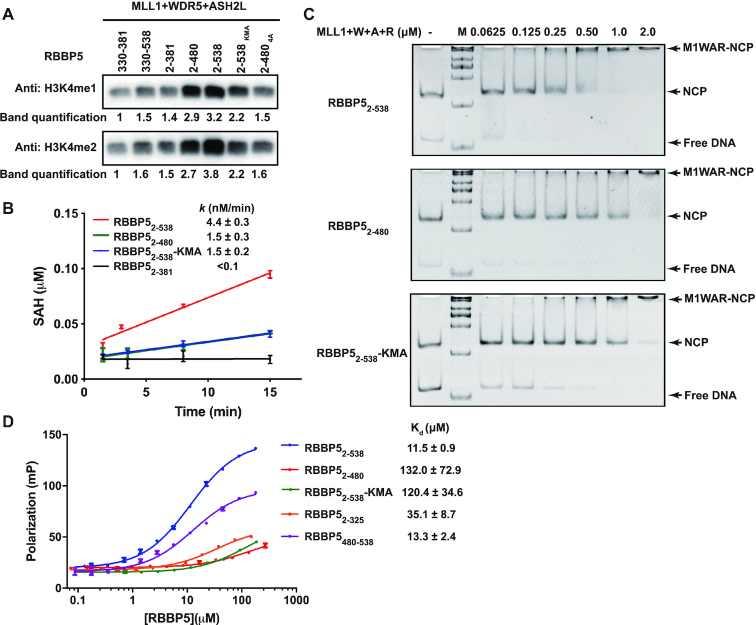
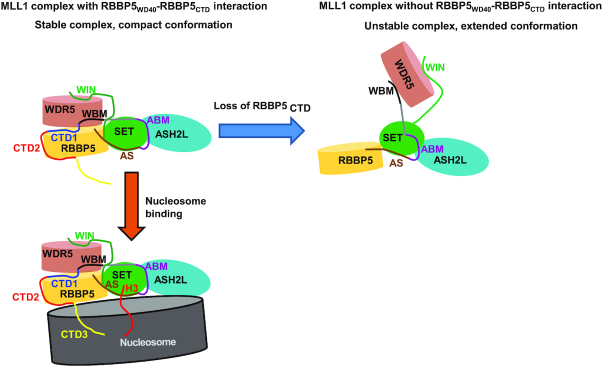
The Mixed Lineage Leukemia protein 1 (MLL1) plays an essential role in the maintenance of the histone H3 lysine 4 (H3K4) methylation status for gene expression during differentiation and development. The methyltransferase activity of MLL1 is regulated by three conserved core subunits, WDR5, RBBP5 and ASH2L. Here, we determined the structure of human RBBP5 and demonstrated its role in the assembly and regulation of the MLL1 complex. We identified an internal interaction between the WD40 propeller and the C-terminal distal region in RBBP5, which assisted the maintenance of the compact conformation of the MLL1 complex. We also discovered a vertebrate-specific motif in the C-terminal distal region of RBBP5 that contributed to nucleosome recognition and methylation of nucleosomes by the MLL1 complex. Our results provide new insights into functional conservation and evolutionary plasticity of the scaffold protein RBBP5 in the regulation of KMT2-family methyltransferase complexes.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Ruthenburg A.J., Allis C.D., Wysocka J.. Methylation of lysine 4 on histone H3: intricacy of writing and reading a single epigenetic mark. Mol. Cell. 2007; 25:15–30. - PubMed
-
- Barski A., Cuddapah S., Cui K., Roh T.Y., Schones D.E., Wang Z., Wei G., Chepelev I., Zhao K.. High-resolution profiling of histone methylations in the human genome. Cell. 2007; 129:823–837. - PubMed
-
- Heintzman N.D., Stuart R.K., Hon G., Fu Y., Ching C.W., Hawkins R.D., Barrera L.O., Van Calcar S., Qu C., Ching K.A. et al.. Distinct and predictive chromatin signatures of transcriptional promoters and enhancers in the human genome. Nat. Genet. 2007; 39:311–318. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
