

Dimensions of control for subthreshold oscillations and spontaneous firing in dopamine neurons
- PMID: 31545787
- PMCID: PMC6776370
- DOI: 10.1371/journal.pcbi.1007375
Dimensions of control for subthreshold oscillations and spontaneous firing in dopamine neurons
Abstract
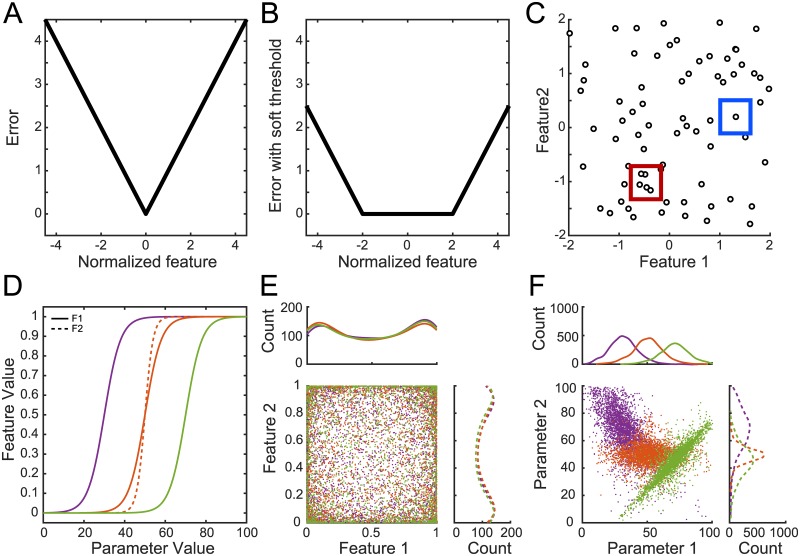
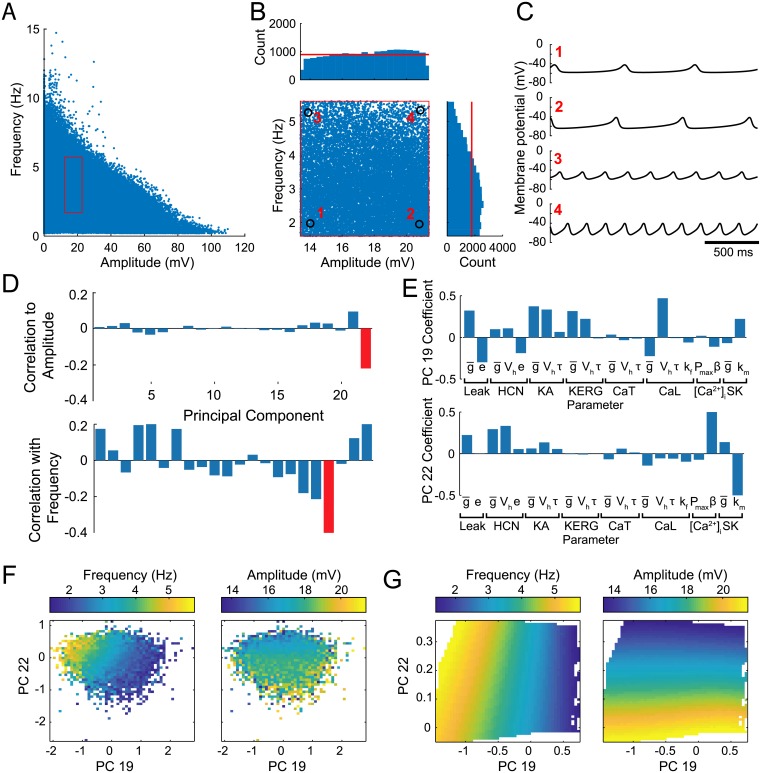
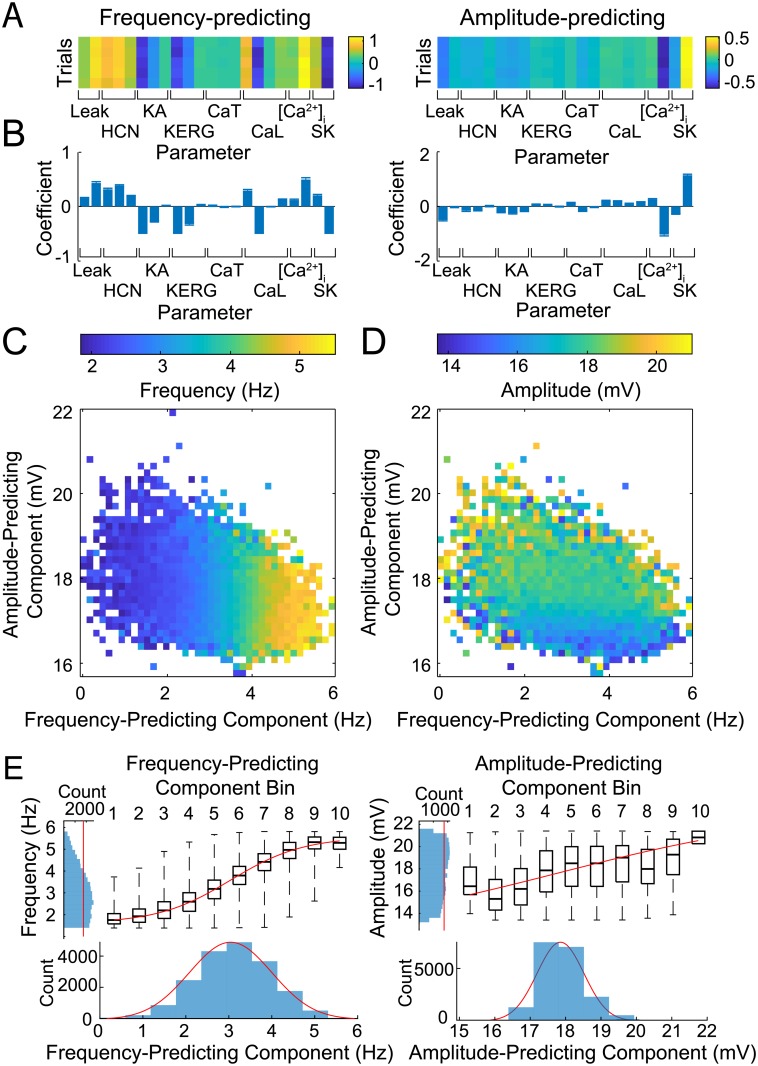
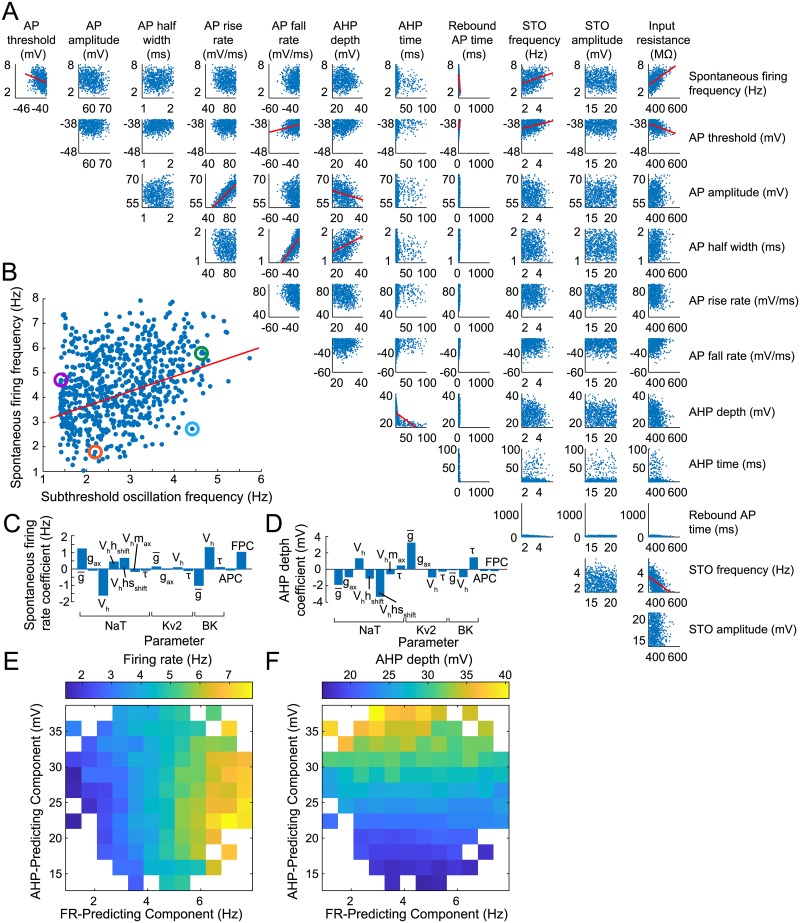
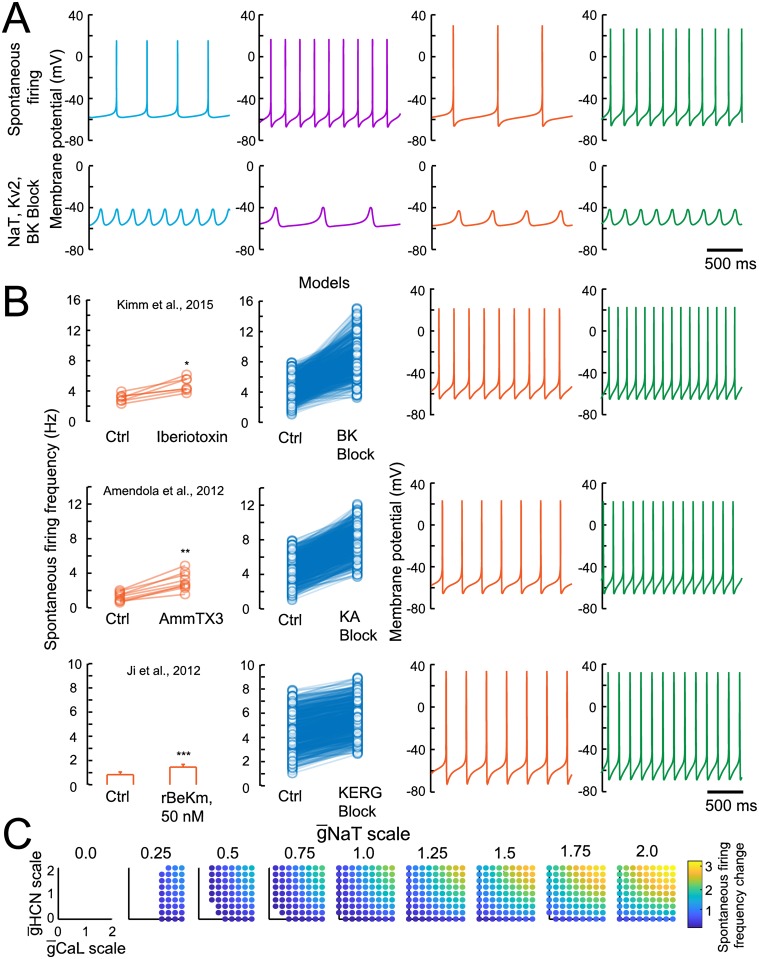
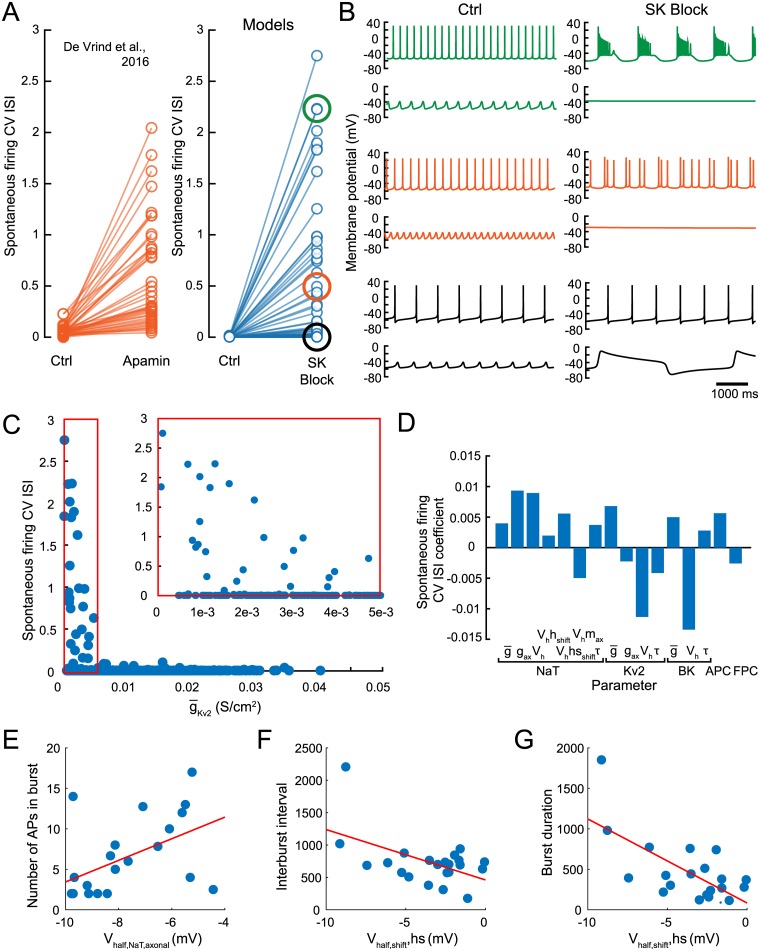
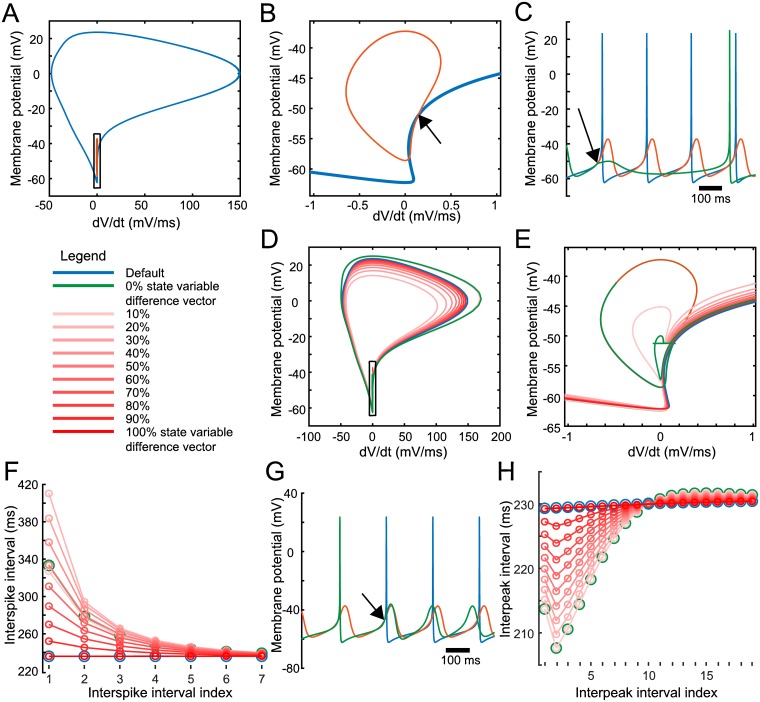
Dopaminergic neurons (DAs) of the rodent substantia nigra pars compacta (SNc) display varied electrophysiological properties in vitro. Despite this, projection patterns and functional inputs from DAs to other structures are conserved, so in vivo delivery of consistent, well-timed dopamine modulation to downstream circuits must be coordinated. Here we show robust coordination by linear parameter controllers, discovered through powerful mathematical analyses of data and models, and from which consistent control of DA subthreshold oscillations (STOs) and spontaneous firing emerges. These units of control represent coordinated intracellular variables, sufficient to regulate complex cellular properties with radical simplicity. Using an evolutionary algorithm and dimensionality reduction, we discovered metaparameters, which when regressed against STO features, revealed a 2-dimensional control plane for the neuron's 22-dimensional parameter space that fully maps the natural range of DA subthreshold electrophysiology. This plane provided a basis for spiking currents to reproduce a large range of the naturally occurring spontaneous firing characteristics of SNc DAs. From it we easily produced a unique population of models, derived using unbiased parameter search, that show good generalization to channel blockade and compensatory intracellular mechanisms. From this population of models, we then discovered low-dimensional controllers for regulating spontaneous firing properties, and gain insight into how currents active in different voltage regimes interact to produce the emergent activity of SNc DAs. Our methods therefore reveal simple regulators of neuronal function lurking in the complexity of combined ion channel dynamics.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: TR and JK are employees of the IBM Corporation.
Figures
Similar articles
-
Calcium dynamics control K-ATP channel-mediated bursting in substantia nigra dopamine neurons: a combined experimental and modeling study.J Neurophysiol. 2018 Jan 1;119(1):84-95. doi: 10.1152/jn.00351.2017. Epub 2017 Oct 4. J Neurophysiol. 2018. PMID: 28978764 Free PMC article.
-
Physiological properties of zebra finch ventral tegmental area and substantia nigra pars compacta neurons.J Neurophysiol. 2006 Nov;96(5):2295-306. doi: 10.1152/jn.01040.2005. Epub 2006 Jul 26. J Neurophysiol. 2006. PMID: 16870835
-
Action potential and calcium dependence of tonic somatodendritic dopamine release in the Substantia Nigra pars compacta.J Neurochem. 2019 Feb;148(4):462-479. doi: 10.1111/jnc.14587. Epub 2018 Nov 12. J Neurochem. 2019. PMID: 30203851
-
Activity-dependent regulation of the dopamine phenotype in substantia nigra neurons.J Neurochem. 2012 May;121(4):497-515. doi: 10.1111/j.1471-4159.2012.07703.x. Epub 2012 Mar 14. J Neurochem. 2012. PMID: 22356203 Review.
-
GABAergic control of substantia nigra dopaminergic neurons.Prog Brain Res. 2007;160:189-208. doi: 10.1016/S0079-6123(06)60011-3. Prog Brain Res. 2007. PMID: 17499115 Review.
Cited by
-
Single Neuron Modeling Identifies Potassium Channel Modulation as Potential Target for Repetitive Head Impacts.Neuroinformatics. 2023 Jul;21(3):501-516. doi: 10.1007/s12021-023-09633-7. Epub 2023 Jun 9. Neuroinformatics. 2023. PMID: 37294503 Free PMC article.
-
Novel and flexible parameter estimation methods for data-consistent inversion in mechanistic modelling.R Soc Open Sci. 2023 Nov 15;10(11):230668. doi: 10.1098/rsos.230668. eCollection 2023 Nov. R Soc Open Sci. 2023. PMID: 38026012 Free PMC article.
-
Enhanced synaptic excitation of VTA dopamine neurons in a mouse model of Alzheimer's disease.bioRxiv [Preprint]. 2025 Jul 31:2025.07.24.666429. doi: 10.1101/2025.07.24.666429. bioRxiv. 2025. PMID: 40766396 Free PMC article. Preprint.
-
An oscillating reaction network with an exact closed form solution in the time domain.BMC Bioinformatics. 2023 Dec 9;24(1):466. doi: 10.1186/s12859-023-05600-w. BMC Bioinformatics. 2023. PMID: 38071308 Free PMC article.
-
Neuronal population models reveal specific linear conductance controllers sufficient to rescue preclinical disease phenotypes.iScience. 2021 Oct 14;24(11):103279. doi: 10.1016/j.isci.2021.103279. eCollection 2021 Nov 19. iScience. 2021. PMID: 34778727 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
