

Human aquaporin-11 guarantees efficient transport of H2O2 across the endoplasmic reticulum membrane
- PMID: 31546170
- PMCID: PMC6812059
- DOI: 10.1016/j.redox.2019.101326
Human aquaporin-11 guarantees efficient transport of H2O2 across the endoplasmic reticulum membrane
Abstract
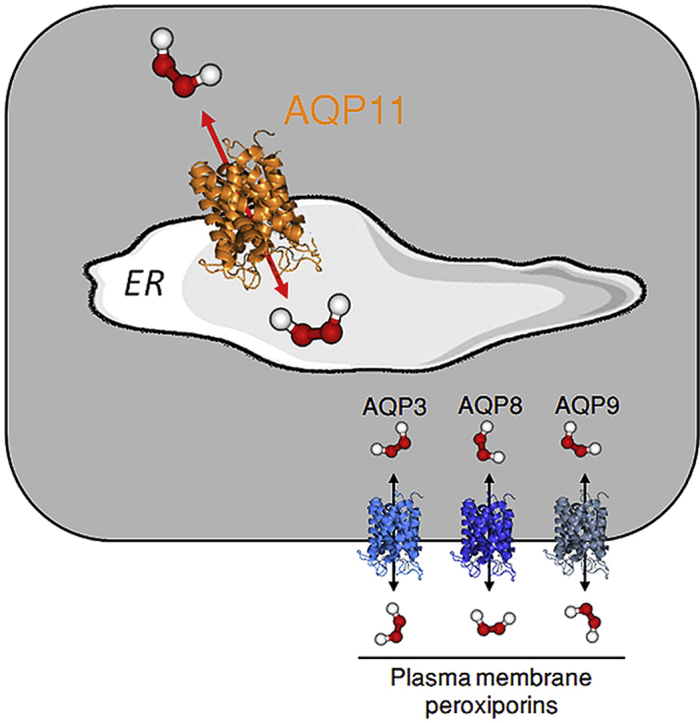
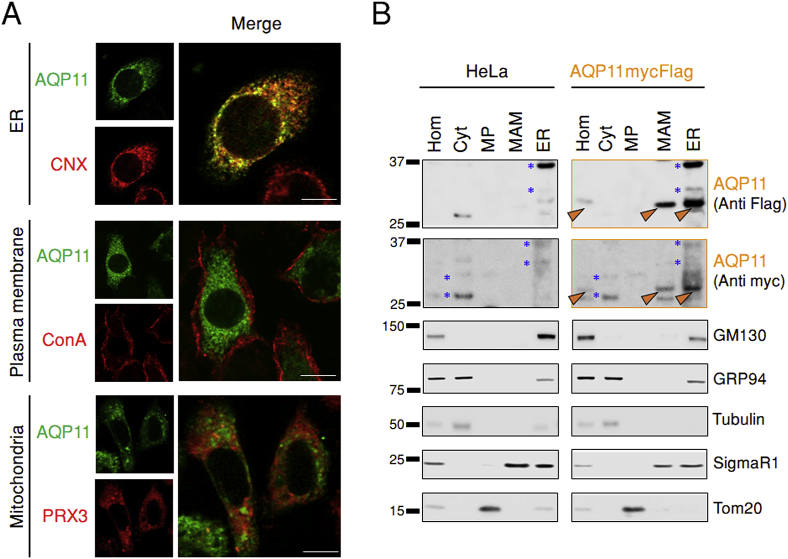
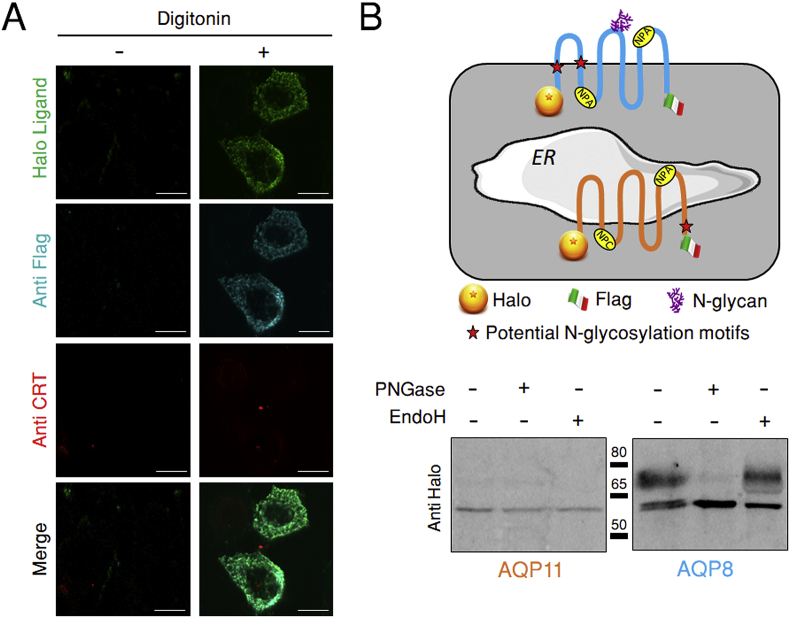
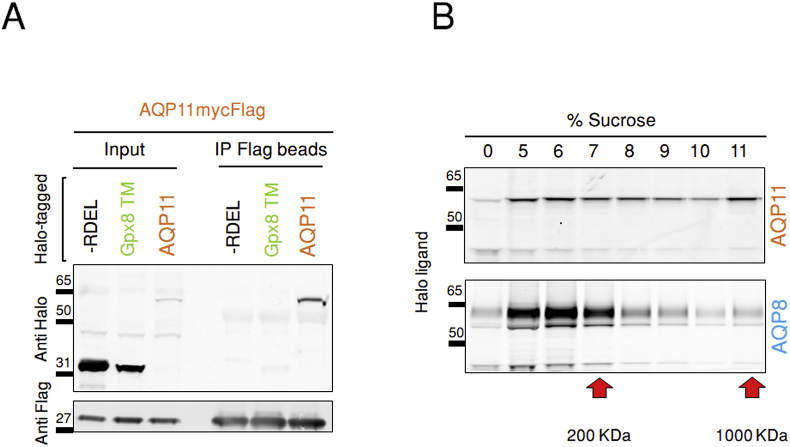
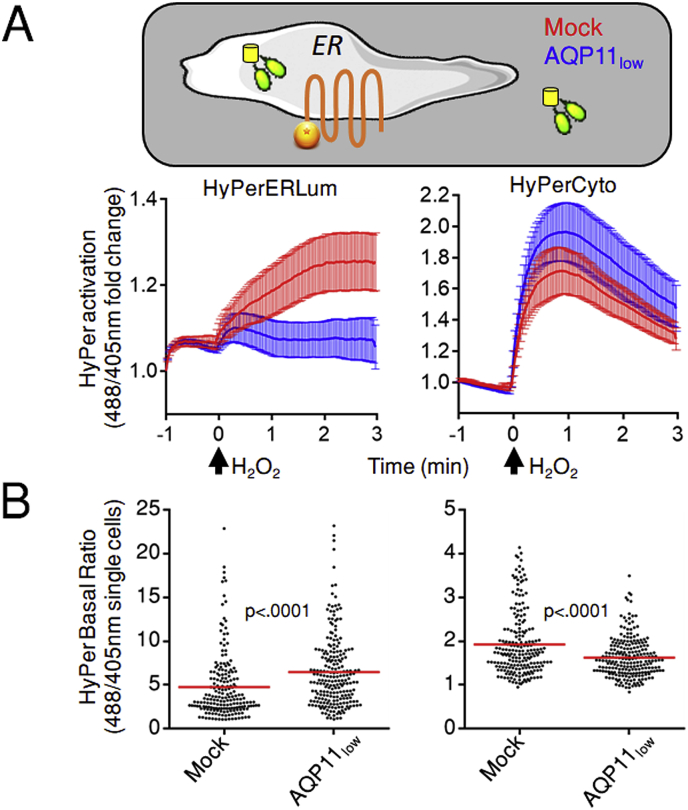
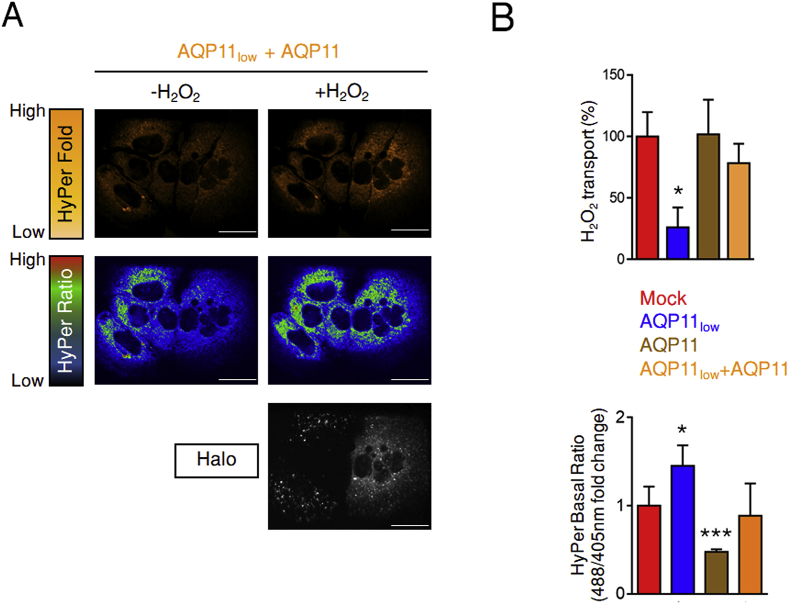
Hydrogen peroxide (H2O2) is an essential second intracellular messenger. To reach its targets in the cytosol, H2O2 must cross a membrane, a feat that requires aquaporins (AQP) endowed with 'peroxiporin' activity (AQP3, AQP8, AQP9). Here, we exploit different organelle-targeted H2O2-sensitive probes to show that also AQP11 efficiently conduits H2O2. Unlike other peroxiporins, AQP11 is localized in the endoplasmic reticulum (ER), accumulating partly in mitochondrial-associated ER membranes (MAM). Its downregulation severely perturbs the flux of H2O2 through the ER, but not through the mitochondrial or plasma membranes. These properties make AQP11 a potential regulator of ER redox homeostasis and signaling.
Keywords: Aquaporins; Endoplasmic reticulum; Hydrogen peroxide; Membrane permeability; Peroxiporins; Redox homeostasis.
Copyright © 2019 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Aquaporin-9 facilitates membrane transport of hydrogen peroxide in mammalian cells.Biochem Biophys Res Commun. 2016 Feb 26;471(1):191-7. doi: 10.1016/j.bbrc.2016.01.153. Epub 2016 Feb 4. Biochem Biophys Res Commun. 2016. PMID: 26837049
-
Transfer of H2O2 from Mitochondria to the endoplasmic reticulum via Aquaporin-11.Redox Biol. 2022 Sep;55:102410. doi: 10.1016/j.redox.2022.102410. Epub 2022 Jul 16. Redox Biol. 2022. PMID: 35863264 Free PMC article.
-
Protective roles of peroxiporins AQP0 and AQP11 in human astrocyte and neuronal cell lines in response to oxidative and inflammatory stressors.Biosci Rep. 2024 Mar 29;44(3):BSR20231725. doi: 10.1042/BSR20231725. Biosci Rep. 2024. PMID: 38451099 Free PMC article.
-
The role of mammalian superaquaporins inside the cell: An update.Biochim Biophys Acta Biomembr. 2021 Jul 1;1863(7):183617. doi: 10.1016/j.bbamem.2021.183617. Epub 2021 Mar 31. Biochim Biophys Acta Biomembr. 2021. PMID: 33811846 Review.
-
Aquaporin-facilitated transmembrane diffusion of hydrogen peroxide.Biochim Biophys Acta. 2014 May;1840(5):1596-604. doi: 10.1016/j.bbagen.2013.09.017. Epub 2013 Sep 20. Biochim Biophys Acta. 2014. PMID: 24060746 Review.
Cited by
-
Hydrogen peroxide in the ER: A tale of triage.Redox Biol. 2020 Jan;28:101358. doi: 10.1016/j.redox.2019.101358. Epub 2019 Oct 22. Redox Biol. 2020. PMID: 31685402 Free PMC article.
-
The Effect of Nutritional Ketosis on Aquaporin Expression in Apolipoprotein E-Deficient Mice: Potential Implications for Energy Homeostasis.Biomedicines. 2022 May 18;10(5):1159. doi: 10.3390/biomedicines10051159. Biomedicines. 2022. PMID: 35625895 Free PMC article.
-
Post-Translational Modification of Cysteines: A Key Determinant of Endoplasmic Reticulum-Mitochondria Contacts (MERCs).Contact (Thousand Oaks). 2021 Mar 24;4:25152564211001213. doi: 10.1177/25152564211001213. eCollection 2021 Jan-Dec. Contact (Thousand Oaks). 2021. PMID: 37366382 Free PMC article. Review.
-
Recent Update on the Molecular Mechanisms of Gonadal Steroids Action in Adipose Tissue.Int J Mol Sci. 2021 May 14;22(10):5226. doi: 10.3390/ijms22105226. Int J Mol Sci. 2021. PMID: 34069293 Free PMC article. Review.
-
Novel Ion Channel Targets and Drug Delivery Tools for Controlling Glioblastoma Cell Invasiveness.Int J Mol Sci. 2021 Nov 2;22(21):11909. doi: 10.3390/ijms222111909. Int J Mol Sci. 2021. PMID: 34769339 Free PMC article. Review.
References
-
- Finn R.N., Cerda J. Evolution and functional diversity of aquaporins. Biol. Bull. 2015;229(1):6–23. - PubMed
-
- Tornroth-Horsefield S., Hedfalk K., Fischer G., Lindkvist-Petersson K., Neutze R. Structural insights into eukaryotic aquaporin regulation. FEBS Lett. 2010;584(12):2580–2588. - PubMed
-
- Murata K., Mitsuoka K., Hirai T., Walz T., Agre P., Heymann J.B. Structural determinants of water permeation through aquaporin-1. Nature. 2000;407(6804):599–605. - PubMed
-
- Verkman A.S. Aquaporins at a glance. J. Cell Sci. 2011;124(13):2107–2112. - PubMed
-
- Ishibashi K., Tanaka Y., Morishita Y. The role of mammalian superaquaporins inside the cell. Biochim. Biophys. Acta. 2014;1840(5):1507–1512. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
