

An Avirulent Strain of Soybean Mosaic Virus Reverses the Defensive Effect of Abscisic Acid in a Susceptible Soybean Cultivar
- PMID: 31546878
- PMCID: PMC6783863
- DOI: 10.3390/v11090879
An Avirulent Strain of Soybean Mosaic Virus Reverses the Defensive Effect of Abscisic Acid in a Susceptible Soybean Cultivar
Abstract
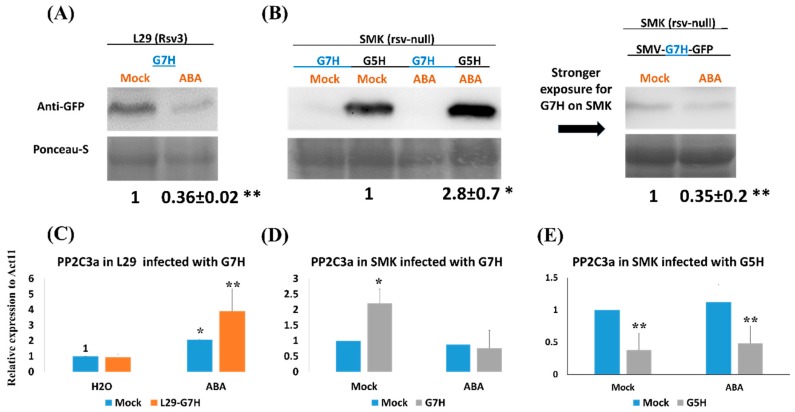
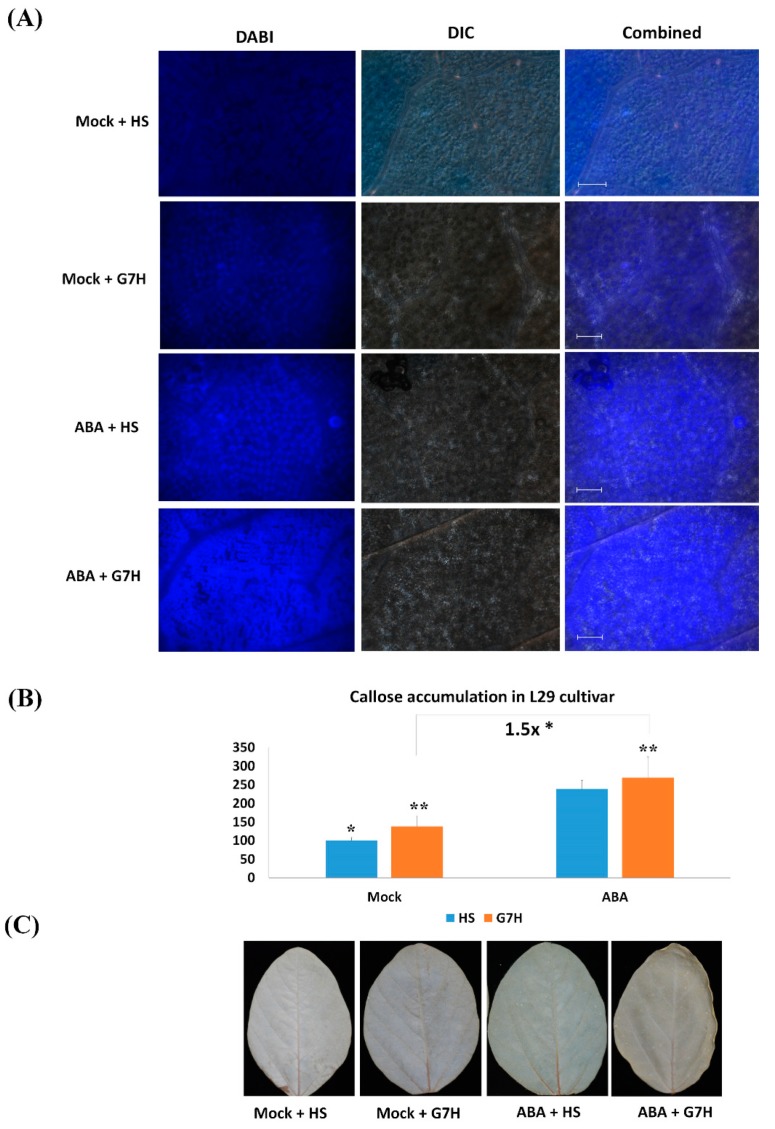
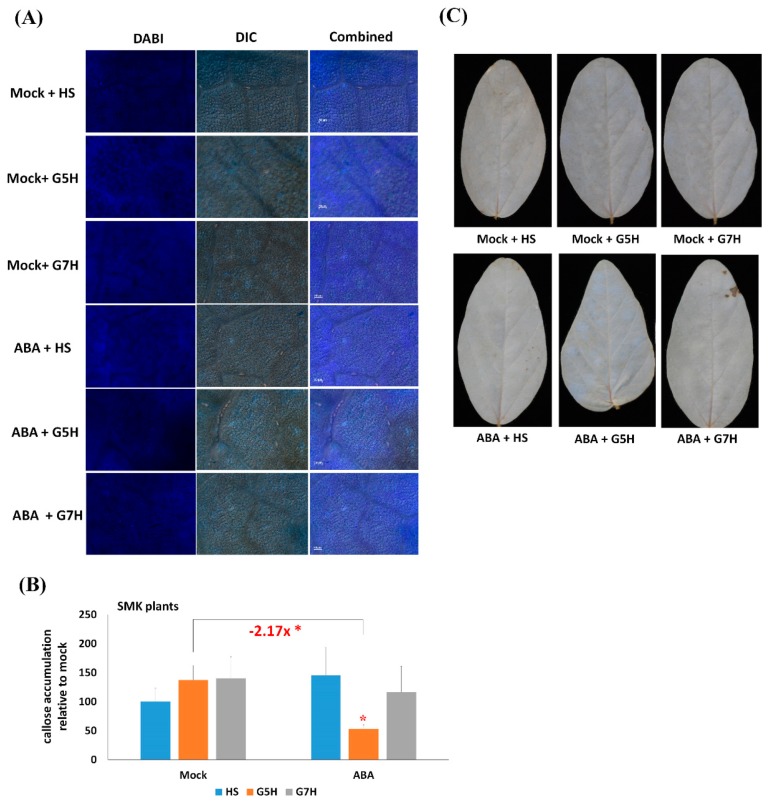
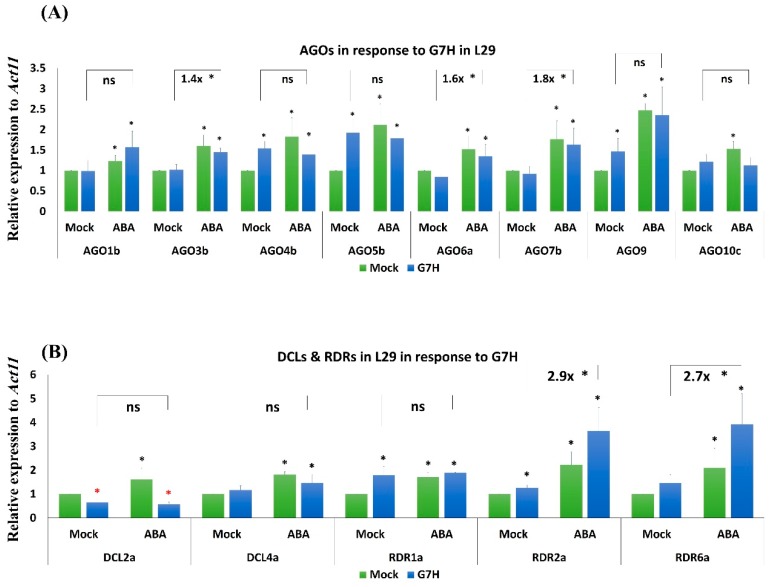
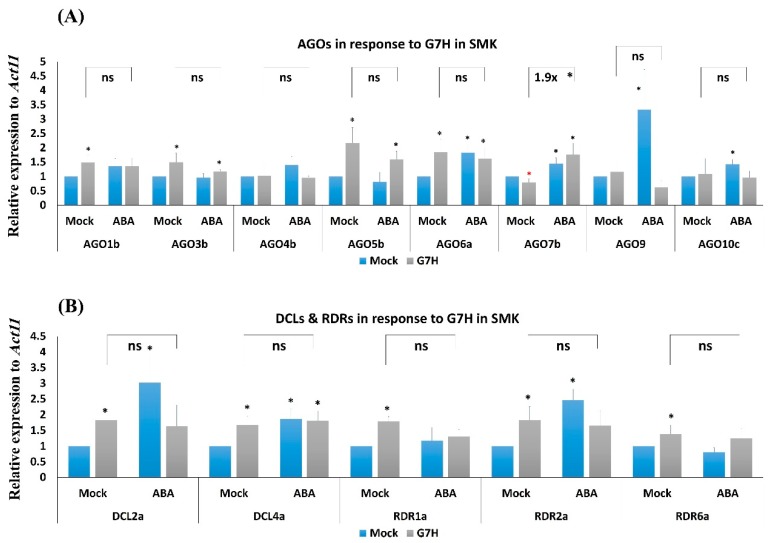
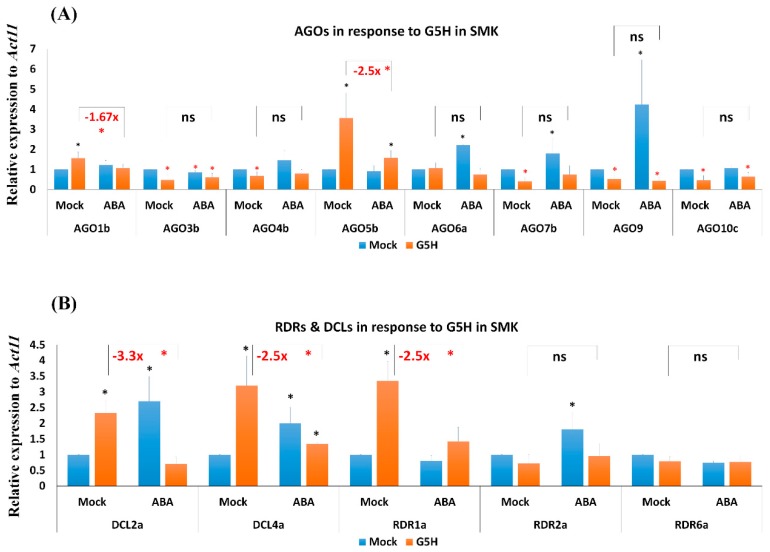
In soybean cultivar L29, the Rsv3 gene is responsible for extreme resistance (ER) against the soybean mosaic virus avirulent strain G5H, but is ineffective against the virulent strain G7H. Part of this ER is attributed to the rapid increase in abscisic acid (ABA) and callose, and to the rapid induction of several genes in the RNA-silencing pathway. Whether these two defense mechanisms are correlated or separated in the ER is unknown. Here, we found that ABA treatment of L29 plants increased the expression of several antiviral RNA-silencing genes as well as the PP2C3a gene, which was previously shown to increase callose accumulation; as a consequence, ABA increased the resistance of L29 plants to G7H. The effect of ABA treatment on these genes was weaker in the rsv3-null cultivar (Somyungkong) than in L29. Besides, G5H-infection of Somyungkong plants subverted the effect of ABA leading to reduced callose accumulation and decreased expression of several RNA-silencing genes, which resulted in increased susceptibility to G5H infection. ABA treatment, however, still induced some resistance to G7H in Somyungkong, but only AGO7b was significantly induced. Our data suggest that Rsv3 modulates the effect of ABA on these two resistance mechanisms, i.e., callose accumulation and the antiviral RNA-silencing pathway, and that in the absence of Rsv3, some strains can reverse the effect of ABA and thereby facilitate their replication and spread.
Keywords: RNA-silencing pathway; Rsv3; abscisic acid; callose; extreme resistance; plant virus; plant–virus interactions; soybean mosaic virus.
Conflict of interest statement
The authors declare no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
References
-
- Klepadlo M., Chen P.Y., Shi A.N., Mason R.E., Korth K.L., Srivastava V., Wu C.J. Two Tightly Linked Genes for Soybean Mosaic Virus Resistance in Soybean. Crop Sci. 2017;57:1844–1853. doi: 10.2135/cropsci2016.05.0290. - DOI
-
- Gunduz I., Buss G.R., Ma G., Chen P., Tolin S.A. Genetic analysis of resistance to soybean mosaic virus in OX670 and harosoy soybean. Crop Sci. 2001;41:1785–1791. doi: 10.2135/cropsci2001.1785. - DOI
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
