

Dual-Specific Protein and Lipid Phosphatase PTEN and Its Biological Functions
- PMID: 31548229
- PMCID: PMC6938656
- DOI: 10.1101/cshperspect.a036301
Dual-Specific Protein and Lipid Phosphatase PTEN and Its Biological Functions
Abstract
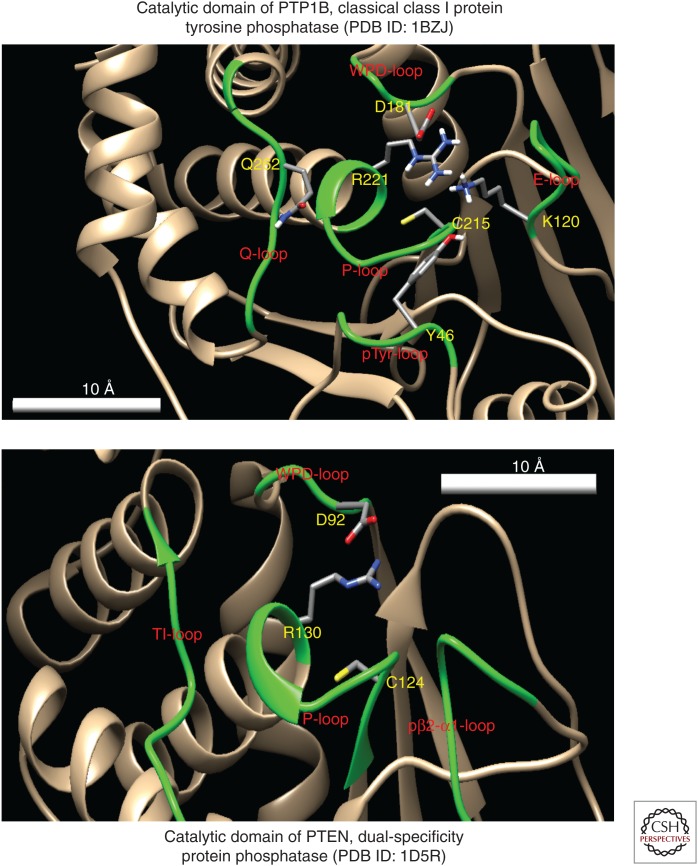
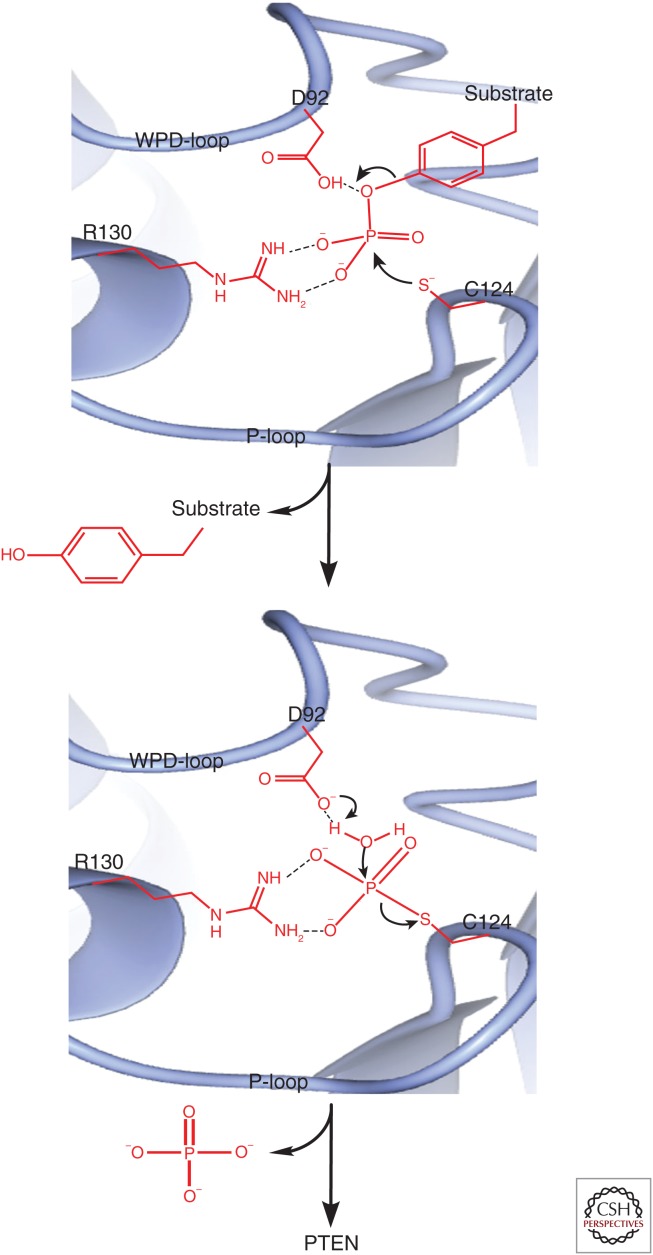
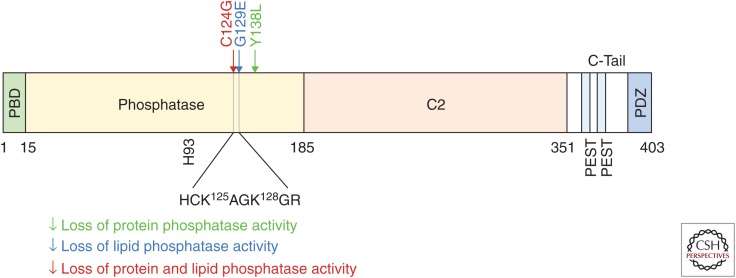
Phosphatase and tensin homolog deleted on chromosome 10 (PTEN) encodes a 403-amino acid protein with an amino-terminal domain that shares sequence homology with the actin-binding protein tensin and the putative tyrosine-protein phosphatase auxilin. Crystal structure analysis of PTEN has revealed a C2 domain that binds to phospholipids in membranes and a phosphatase domain that displays dual-specific activity toward both tyrosine (Y), serine (S)/threonine (T), as well as lipid substrates in vitro. Characterized primarily as a lipid phosphatase, PTEN plays important roles in multiple cellular processes including cell growth/survival as well as metabolism.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
Similar articles
-
Acute regulation of the tumour suppressor phosphatase, PTEN, by anionic lipids and reactive oxygen species.Biochem Soc Trans. 2004 Apr;32(Pt 2):338-42. doi: 10.1042/bst0320338. Biochem Soc Trans. 2004. PMID: 15046604 Review.
-
Analysis of the cellular functions of PTEN using catalytic domain and C-terminal mutations: differential effects of C-terminal deletion on signalling pathways downstream of phosphoinositide 3-kinase.Biochem J. 2000 Mar 15;346 Pt 3(Pt 3):827-33. Biochem J. 2000. PMID: 10698713 Free PMC article.
-
Functional analysis of the protein phosphatase activity of PTEN.Biochem J. 2012 Jun 15;444(3):457-64. doi: 10.1042/BJ20120098. Biochem J. 2012. PMID: 22413754 Free PMC article.
-
Antagonism of PI 3-kinase-dependent signalling pathways by the tumour suppressor protein, PTEN.Biochem Soc Trans. 2001 Nov;29(Pt 6):846-51. doi: 10.1042/0300-5127:0290846. Biochem Soc Trans. 2001. PMID: 11709086 Review.
-
Cell cycle arrest and apoptosis by expression of a novel TPIP (TPIP-C2) cDNA encoding a C2-domain in HEK-293 cells.Mol Biol Rep. 2012 Jul;39(7):7389-402. doi: 10.1007/s11033-012-1571-6. Epub 2012 Feb 7. Mol Biol Rep. 2012. PMID: 22311048
Cited by
-
Chronic Exposure to Palmitic Acid Down-Regulates AKT in Beta-Cells through Activation of mTOR.Am J Pathol. 2022 Jan;192(1):130-145. doi: 10.1016/j.ajpath.2021.09.008. Epub 2021 Oct 5. Am J Pathol. 2022. PMID: 34619135 Free PMC article.
-
Targeting the E3 ligase NEDD4 as a novel therapeutic strategy for IGF1 signal pathway-driven gastric cancer.Oncogene. 2023 Mar;42(14):1072-1087. doi: 10.1038/s41388-023-02619-4. Epub 2023 Feb 11. Oncogene. 2023. PMID: 36774408 Free PMC article.
-
PTEN inhibits AMPK to control collective migration.Nat Commun. 2022 Aug 11;13(1):4528. doi: 10.1038/s41467-022-31842-y. Nat Commun. 2022. PMID: 35953476 Free PMC article.
-
Involvement of the Mitochondrial Protein Tyrosine Phosphatase PTPM1 in the Promotion of Conidiation, Development, and Pathogenicity in Colletotrichum graminicola.Front Microbiol. 2021 Jan 14;11:605738. doi: 10.3389/fmicb.2020.605738. eCollection 2020. Front Microbiol. 2021. PMID: 33519752 Free PMC article.
-
Cancer and Autism: How PTEN Mutations Degrade Function at the Membrane and Isoform Expression in the Human Brain.J Mol Biol. 2023 Dec 15;435(24):168354. doi: 10.1016/j.jmb.2023.168354. Epub 2023 Nov 5. J Mol Biol. 2023. PMID: 37935253 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous