

RNA polymerases as moving barriers to condensin loop extrusion
- PMID: 31548377
- PMCID: PMC6789630
- DOI: 10.1073/pnas.1907009116
RNA polymerases as moving barriers to condensin loop extrusion
Abstract
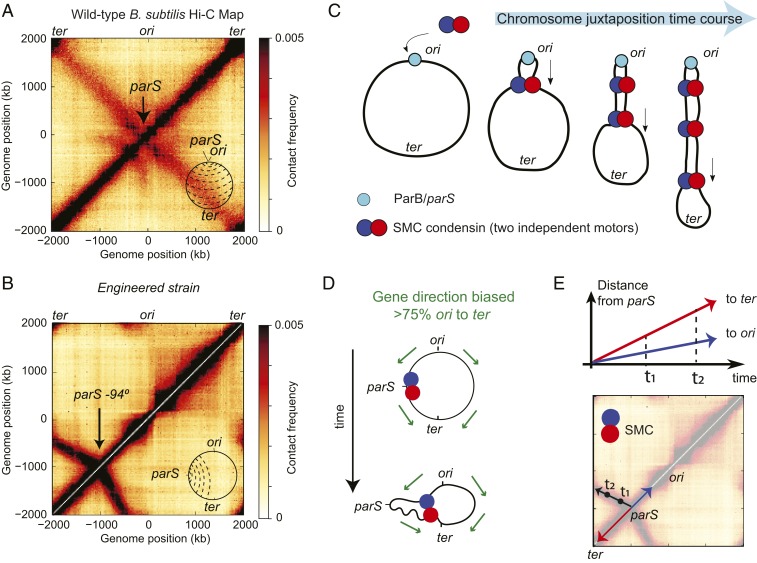
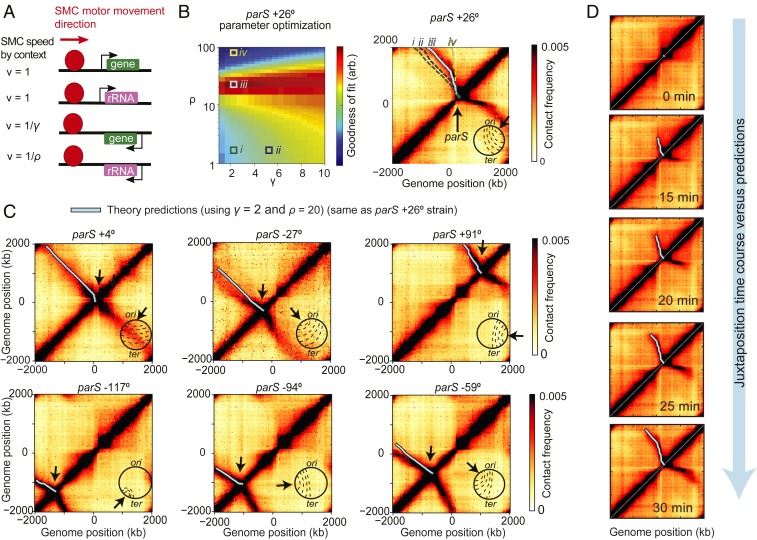
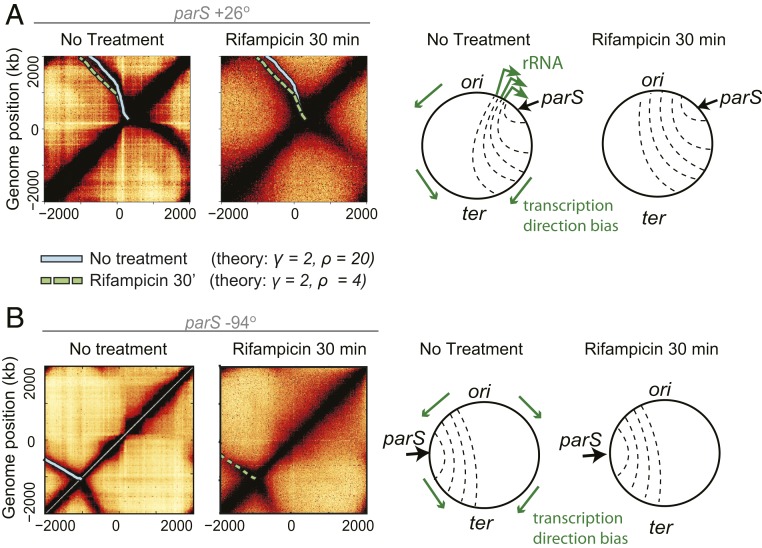
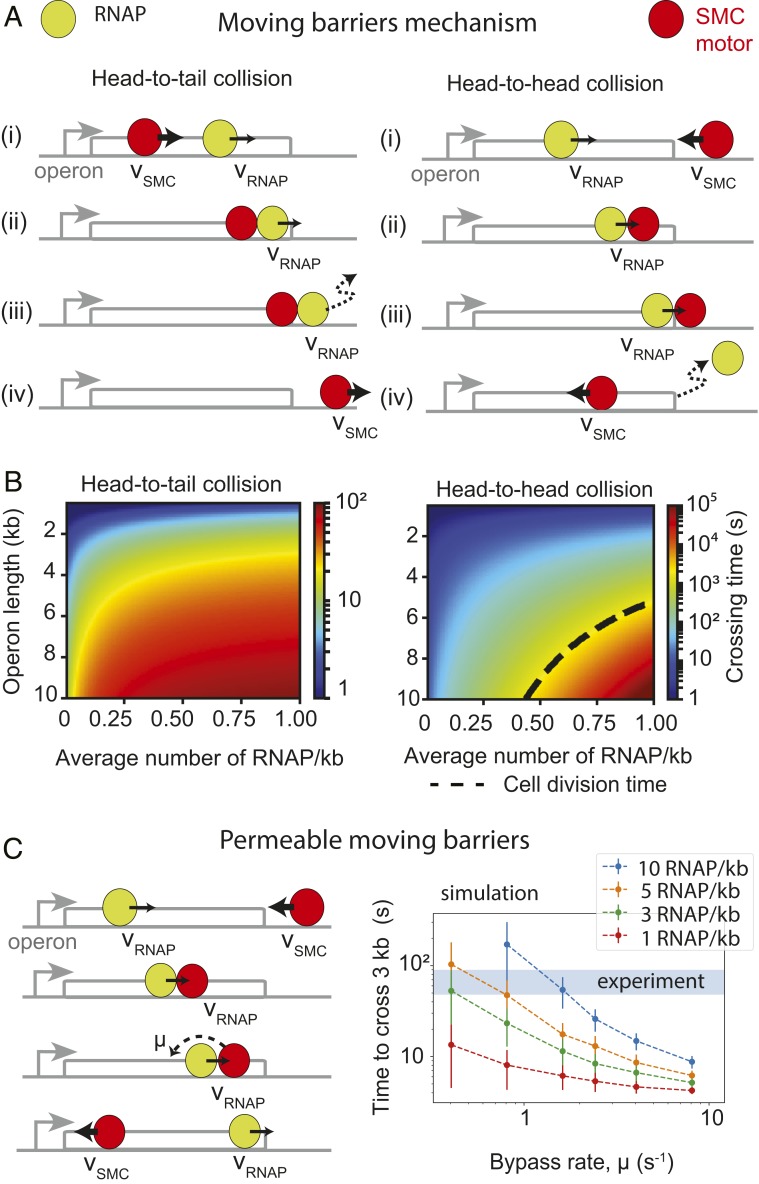
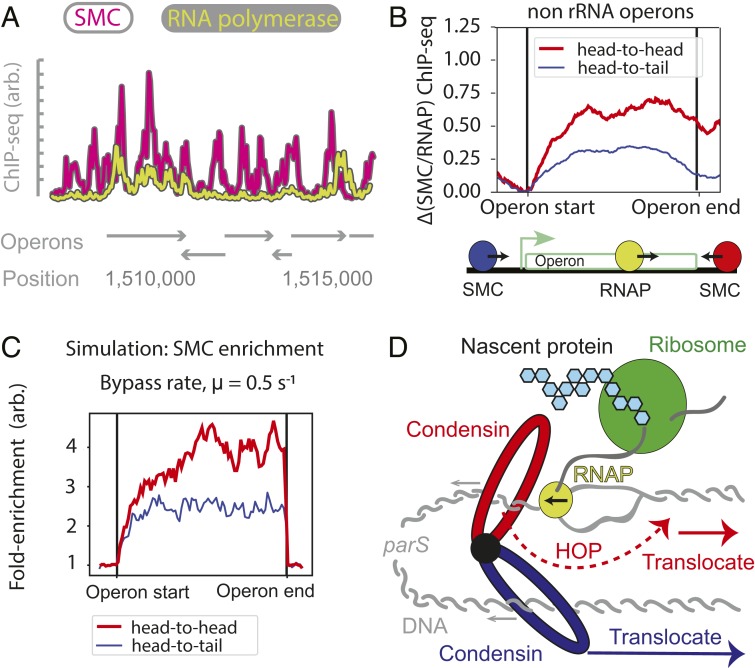
To separate replicated sister chromatids during mitosis, eukaryotes and prokaryotes have structural maintenance of chromosome (SMC) condensin complexes that were recently shown to organize chromosomes by a process known as DNA loop extrusion. In rapidly dividing bacterial cells, the process of separating sister chromatids occurs concomitantly with ongoing transcription. How transcription interferes with the condensin loop-extrusion process is largely unexplored, but recent experiments have shown that sites of high transcription may directionally affect condensin loop extrusion. We quantitatively investigate different mechanisms of interaction between condensin and elongating RNA polymerases (RNAPs) and find that RNAPs are likely steric barriers that can push and interact with condensins. Supported by chromosome conformation capture and chromatin immunoprecipitation for cells after transcription inhibition and RNAP degradation, we argue that translocating condensins must bypass transcribing RNAPs within ∼1 to 2 s of an encounter at rRNA genes and within ∼10 s at protein-coding genes. Thus, while individual RNAPs have little effect on the progress of loop extrusion, long, highly transcribed operons can significantly impede the extrusion process. Our data and quantitative models further suggest that bacterial condensin loop extrusion occurs by 2 independent, uncoupled motor activities; the motors translocate on DNA in opposing directions and function together to enlarge chromosomal loops, each independently bypassing steric barriers in their path. Our study provides a quantitative link between transcription and 3D genome organization and proposes a mechanism of interactions between SMC complexes and elongating transcription machinery relevant from bacteria to higher eukaryotes.
Keywords: SMC; chromosome conformation capture; condensin; loop extrusion; transcription.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hirano T., Condensin-based chromosome organization from bacteria to vertebrates. Cell 164, 847–857 (2016). - PubMed
-
- Riggs A. D., DNA methylation and late replication probably aid cell memory, and type I DNA reeling could aid chromosome folding and enhancer function. Philos. Trans. R. Soc. Lond. B Biol. Sci. 326, 285–297 (1990). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
