

PECTIN ACETYLESTERASE9 Affects the Transcriptome and Metabolome and Delays Aphid Feeding
- PMID: 31551361
- PMCID: PMC6878011
- DOI: 10.1104/pp.19.00635
PECTIN ACETYLESTERASE9 Affects the Transcriptome and Metabolome and Delays Aphid Feeding
Abstract
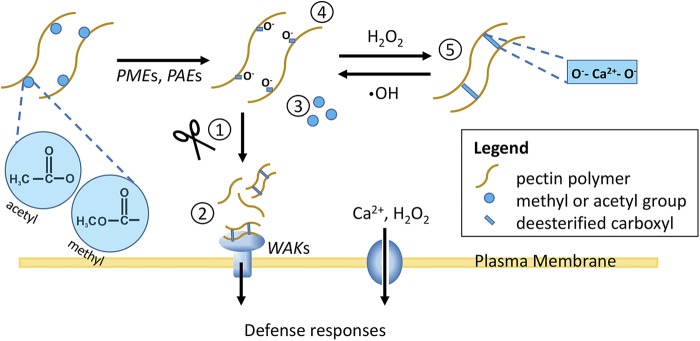
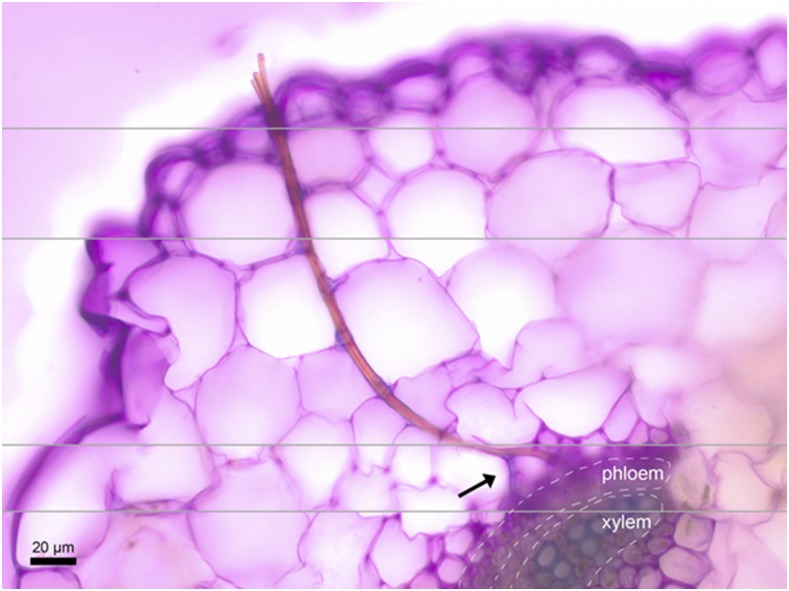
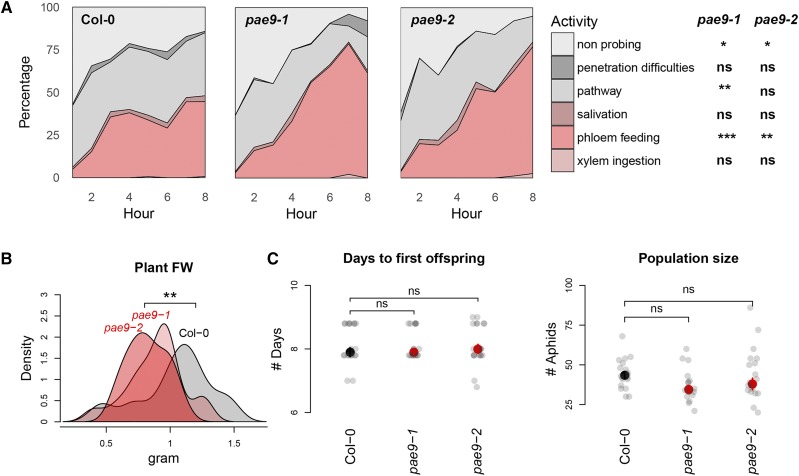
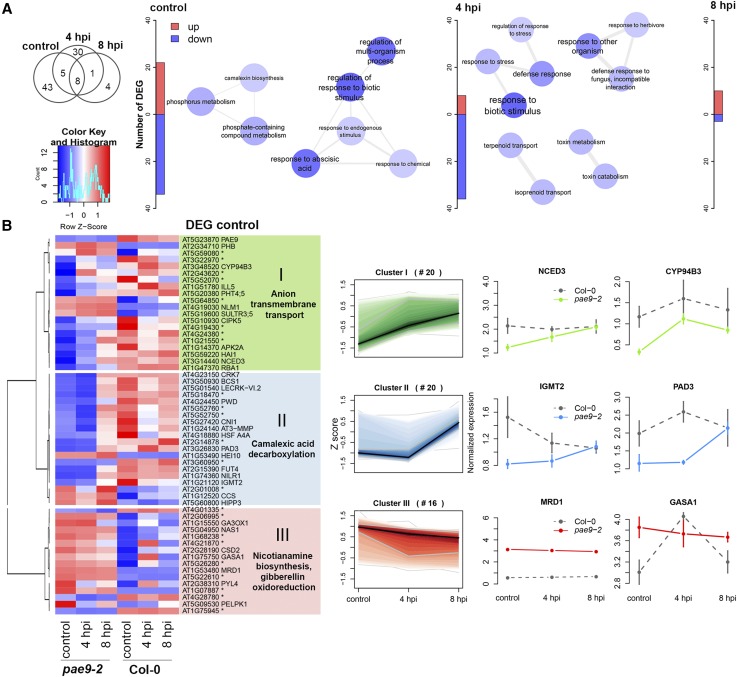
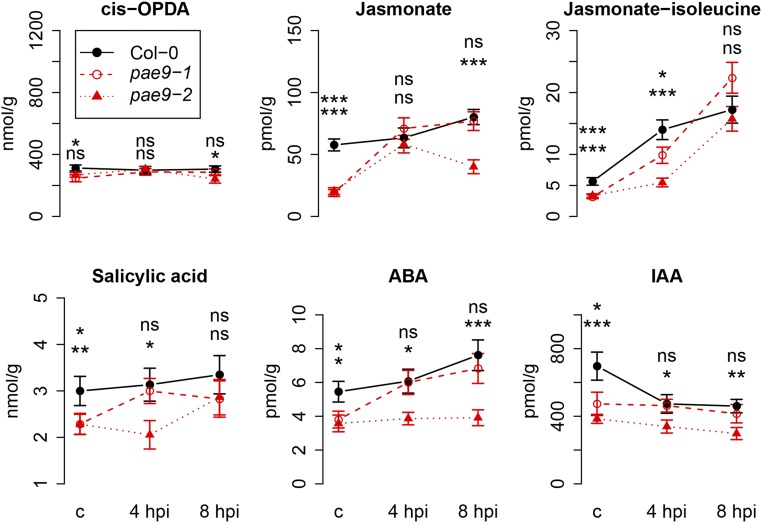
The plant cell wall plays an important role in damage-associated molecular pattern-induced resistance to pathogens and herbivorous insects. Our current understanding of cell wall-mediated resistance is largely based on the degree of pectin methylesterification. However, little is known about the role of pectin acetylesterification in plant immunity. This study describes how one pectin-modifying enzyme, PECTIN ACETYLESTERASE 9 (PAE9), affects the Arabidopsis (Arabidopsis thaliana) transcriptome, secondary metabolome, and aphid performance. Electro-penetration graphs showed that Myzus persicae aphids established phloem feeding earlier on pae9 mutants. Whole-genome transcriptome analysis revealed a set of 56 differentially expressed genes (DEGs) between uninfested pae9-2 mutants and wild-type plants. The majority of the DEGs were enriched for biotic stress responses and down-regulated in the pae9-2 mutant, including PAD3 and IGMT2, involved in camalexin and indole glucosinolate biosynthesis, respectively. Relative quantification of more than 100 secondary metabolites revealed decreased levels of several compounds, including camalexin and oxylipins, in two independent pae9 mutants. In addition, absolute quantification of phytohormones showed that jasmonic acid (JA), jasmonoyl-Ile, salicylic acid, abscisic acid, and indole-3-acetic acid were compromised due to PAE9 loss of function. After aphid infestation, however, pae9 mutants increased their levels of camalexin, glucosinolates, and JA, and no long-term effects were observed on aphid fitness. Overall, these data show that PAE9 is required for constitutive up-regulation of defense-related compounds, but that it is not required for aphid-induced defenses. The signatures of phenolic antioxidants, phytoprostanes, and oxidative stress-related transcripts indicate that the processes underlying PAE9 activity involve oxidation-reduction reactions.
© 2019 American Society of Plant Biologists. All Rights Reserved.
Figures

References
-
- Abdel-Ghany SE, Burkhead JL, Gogolin KA, Andrés-Colás N, Bodecker JR, Puig S, Peñarrubia L, Pilon M (2005) AtCCS is a functional homolog of the yeast copper chaperone Ccs1/Lys7. FEBS Lett 579: 2307–2312 - PubMed
-
- Albersheim P, Jones TM, English PD (1969) Biochemistry of the cell wall in relation to infective processes. Annu Rev Phytopathol 7: 171–194 - PubMed
-
- Aldington S, McDougall GJ, Fry SC (1991) Structure‐activity relationships of biologically active oligosaccharides. Plant Cell Environ 14: 625–636
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
