

Bach2 Deficiency Leads to Spontaneous Expansion of IL-4-Producing T Follicular Helper Cells and Autoimmunity
- PMID: 31552021
- PMCID: PMC6737000
- DOI: 10.3389/fimmu.2019.02050
Bach2 Deficiency Leads to Spontaneous Expansion of IL-4-Producing T Follicular Helper Cells and Autoimmunity
Abstract
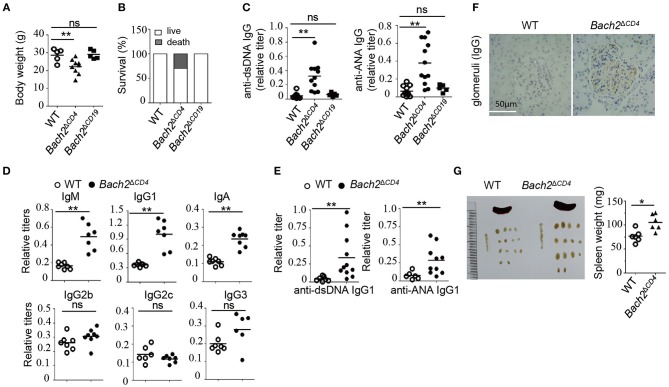
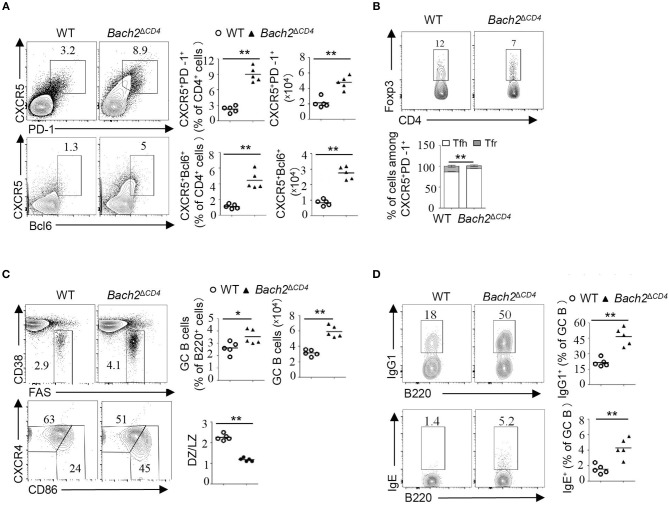
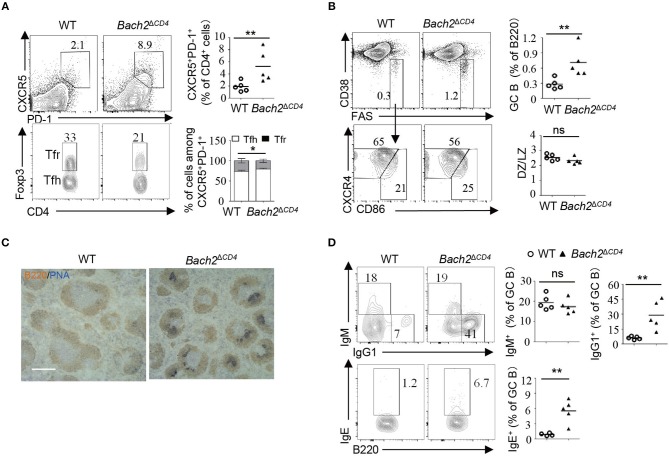
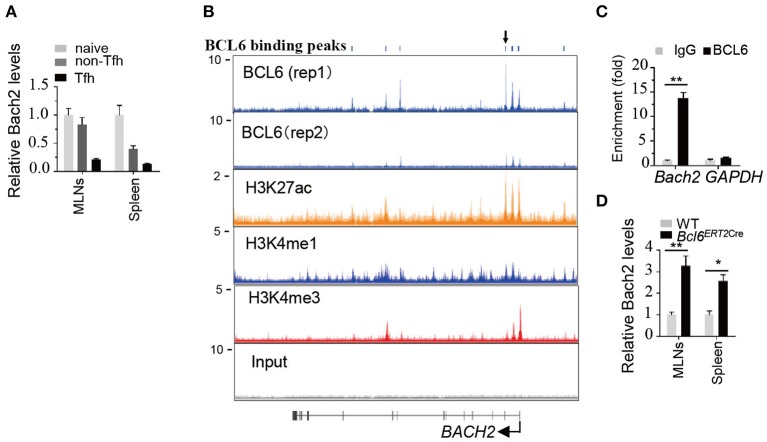
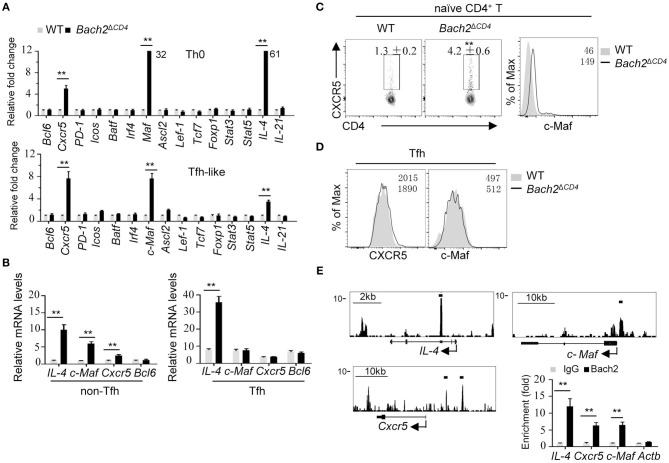
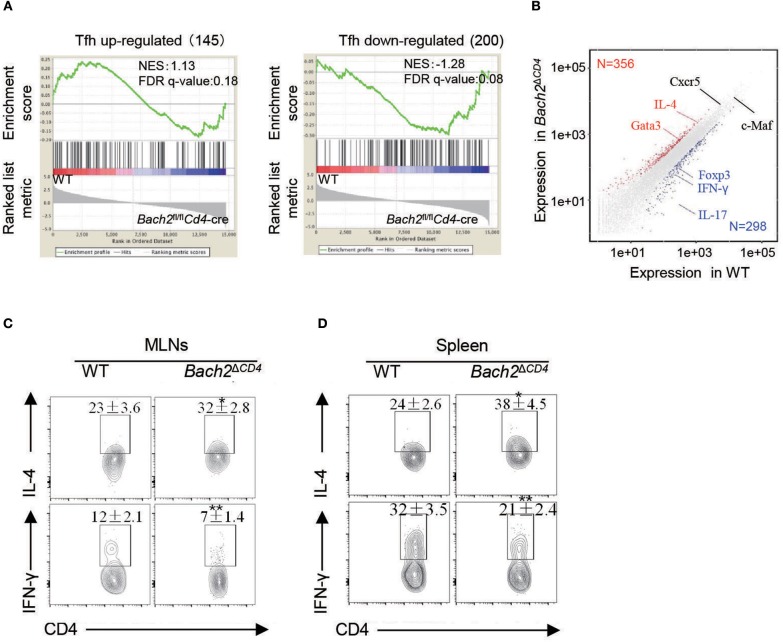
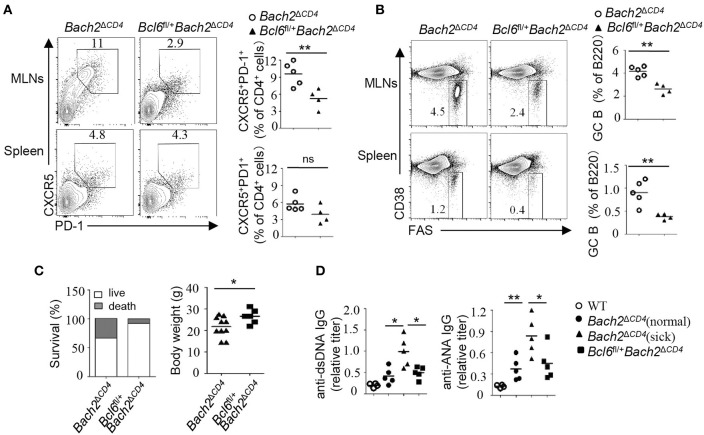
The transcription factor Bach2 is a susceptible gene for numerous autoimmune diseases including systemic lupus erythematosus (SLE). Bach2-/- mice can develop a lupus-like autoimmune disease. However, the exact cellular and molecular mechanisms via which Bach2 protects the hosts from developing autoimmunity remains incompletely understood. Here, we report that Bach2 ablation on T cells, but not B cells, resulted in humoral autoimmunity, and this was associated with expansion of T follicular helper (Tfh) cells and abnormal germinal centers. Bach2 was down-regulated in Tfh cells and directly suppressed by the Tfh-defining transcription factor BCL6. Mechanistically, Bach2 directly suppresses the transcription of Cxcr5 and c-Maf, two key regulators of Tfh cell differentiation. Bach2-deficient Tfh cells were skewed toward the IL-4-producing subset, which induced IgG1 and IgE isotype switching of B cells. Heterozygous Bcl6 deficiency reduced the formation of germinal center and autoantibodies, and ameliorated the pathology in Bach2-deficient mice. Our findings identify Bach2 as a crucial negative regulator of Tfh cells at steady state and prove that Bach2 controls autoimmunity in part by restraining accumulation of pathogenic Tfh cells.
Keywords: BCL6; Bach2; IL-4; T follicular helper cells; autoimmunity.
Figures
Similar articles
-
Bach2 Negatively Regulates T Follicular Helper Cell Differentiation and Is Critical for CD4+ T Cell Memory.J Immunol. 2019 May 15;202(10):2991-2998. doi: 10.4049/jimmunol.1801626. Epub 2019 Apr 10. J Immunol. 2019. PMID: 30971440 Free PMC article.
-
B cell-derived IL-6 initiates spontaneous germinal center formation during systemic autoimmunity.J Exp Med. 2017 Nov 6;214(11):3207-3217. doi: 10.1084/jem.20170580. Epub 2017 Sep 12. J Exp Med. 2017. PMID: 28899868 Free PMC article.
-
The Transcription Factor Ets1 Suppresses T Follicular Helper Type 2 Cell Differentiation to Halt the Onset of Systemic Lupus Erythematosus.Immunity. 2018 Dec 18;49(6):1034-1048.e8. doi: 10.1016/j.immuni.2018.10.012. Immunity. 2018. PMID: 30566881
-
Follicular Helper T Cells in Systemic Lupus Erythematosus.Front Immunol. 2018 Aug 3;9:1793. doi: 10.3389/fimmu.2018.01793. eCollection 2018. Front Immunol. 2018. PMID: 30123218 Free PMC article. Review.
-
Transcriptional regulation of follicular T-helper (Tfh) cells.Immunol Rev. 2013 Mar;252(1):139-45. doi: 10.1111/imr.12040. Immunol Rev. 2013. PMID: 23405901 Free PMC article. Review.
Cited by
-
Stepwise Tfh cell differentiation revisited: new advances and long-standing questions.Fac Rev. 2021;10:3. doi: 10.12703/r/10-3. Epub 2021 Jan 15. Fac Rev. 2021. PMID: 33644779 Free PMC article.
-
SNP-to-gene linking strategies reveal contributions of enhancer-related and candidate master-regulator genes to autoimmune disease.Cell Genom. 2022 Jul 13;2(7):100145. doi: 10.1016/j.xgen.2022.100145. Cell Genom. 2022. PMID: 35873673 Free PMC article.
-
Characterization of rare spindle and root cell transcriptional profiles in the stria vascularis of the adult mouse cochlea.Sci Rep. 2020 Oct 22;10(1):18100. doi: 10.1038/s41598-020-75238-8. Sci Rep. 2020. PMID: 33093630 Free PMC article.
-
Altered Phenotypes of Colonic and Peripheral Blood Follicular Helper and Follicular Cytotoxic T Cells in Mice with DSS-Induced Colitis.J Inflamm Res. 2023 Jul 10;16:2879-2892. doi: 10.2147/JIR.S411373. eCollection 2023. J Inflamm Res. 2023. PMID: 37456782 Free PMC article.
-
PD-L1- and IL-4-expressing basophils promote pathogenic accumulation of T follicular helper cells in lupus.Nat Commun. 2024 Apr 22;15(1):3389. doi: 10.1038/s41467-024-47691-w. Nat Commun. 2024. PMID: 38649353 Free PMC article.
References
-
- International Multiple Sclerosis Genetics Consortium, Wellcome Trust Case Control Consortium. Sawcer S, Hellenthal G, Pirinen M, Spencer CC, et al. . Genetic risk and a primary role for cell-mediated immune mechanisms in multiple sclerosis. Nature. (2011) 476:214–9. 10.1038/nature10251 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases