

Low temperature traps myosin motors of mammalian muscle in a refractory state that prevents activation
- PMID: 31554652
- PMCID: PMC6829559
- DOI: 10.1085/jgp.201912424
Low temperature traps myosin motors of mammalian muscle in a refractory state that prevents activation
Abstract
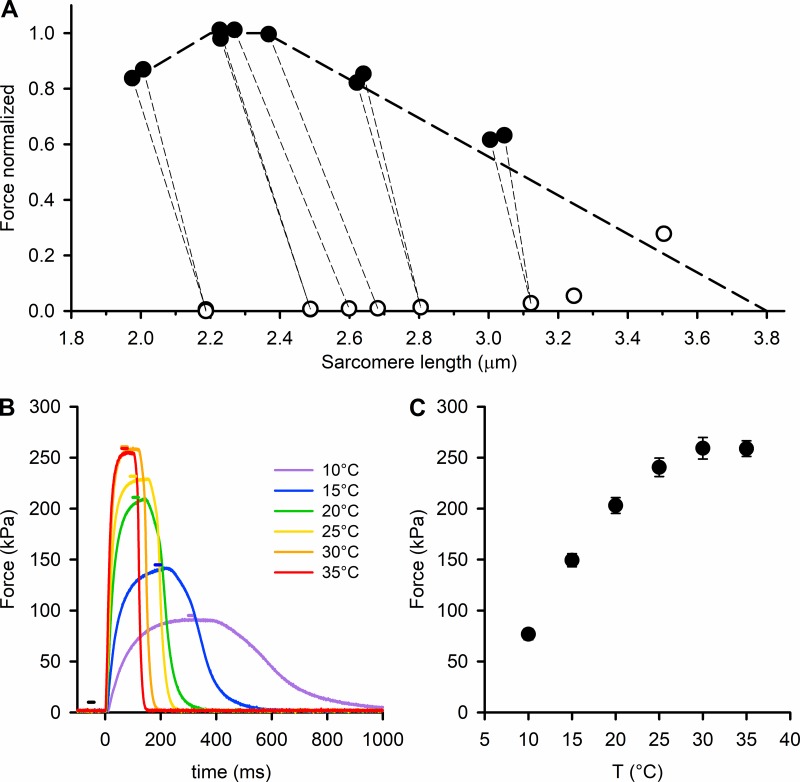
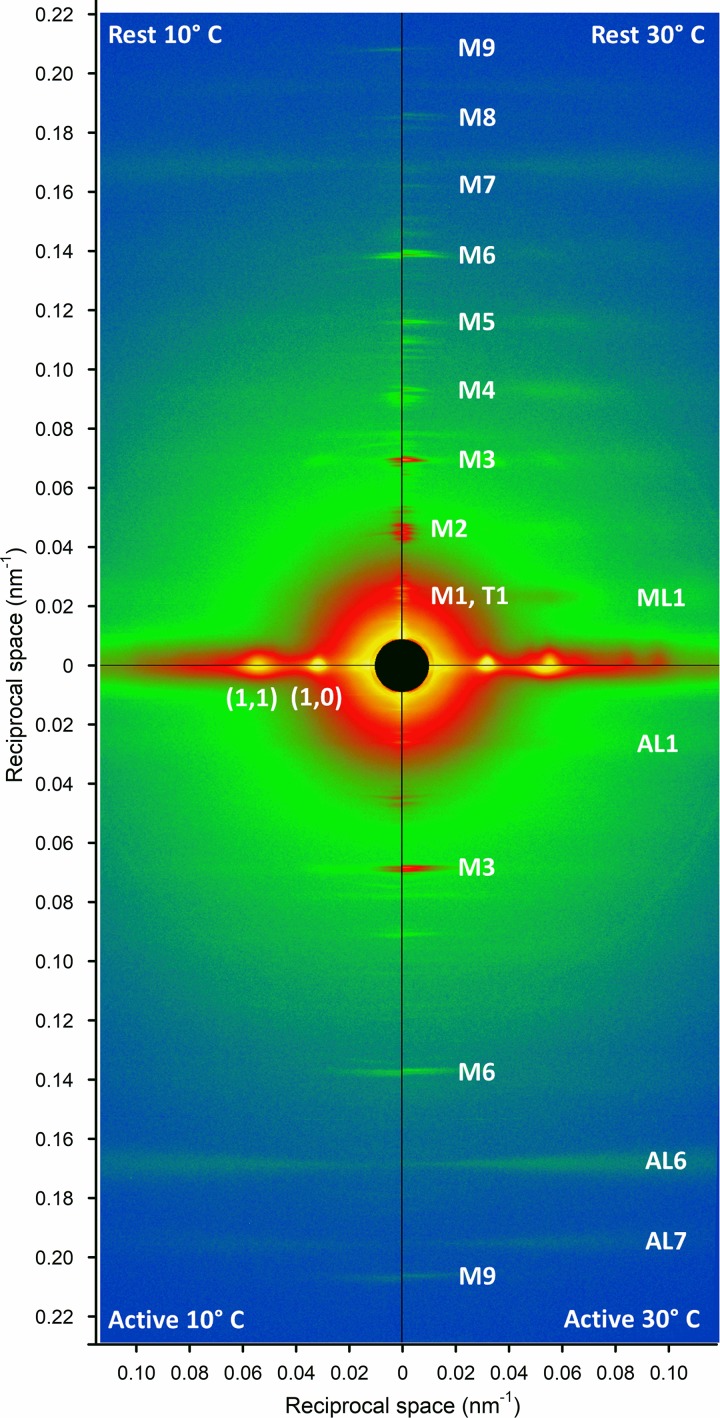
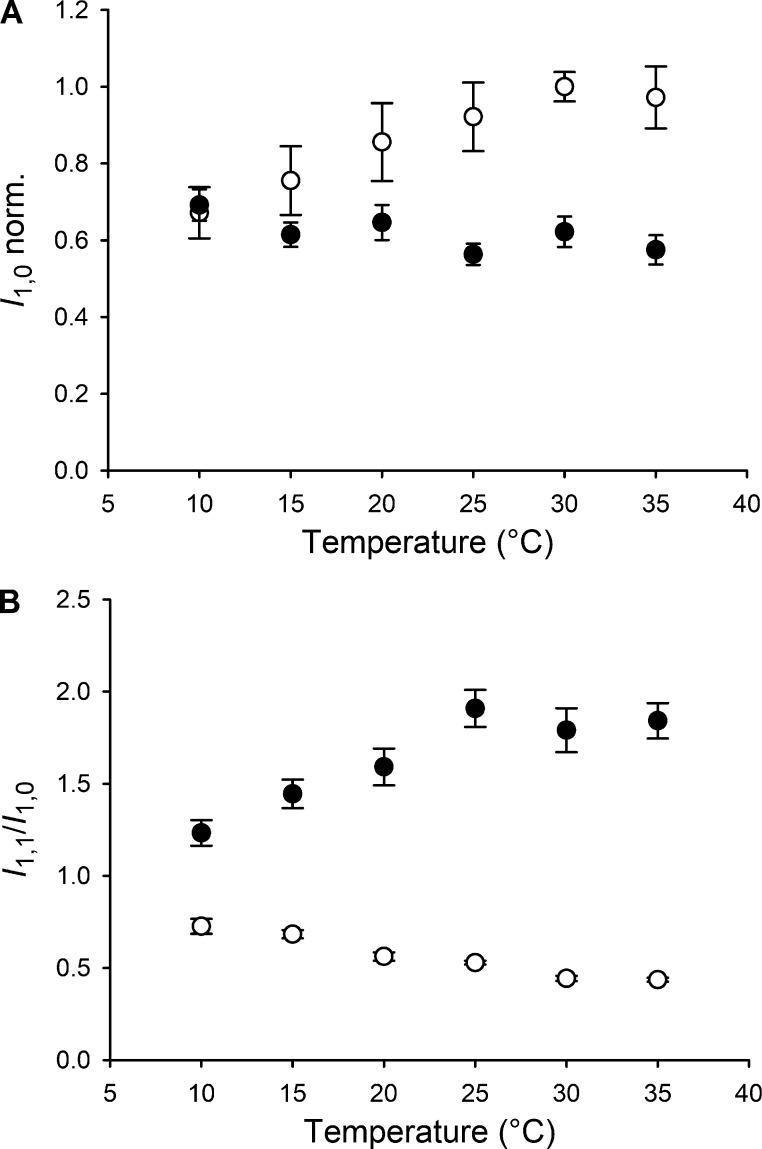
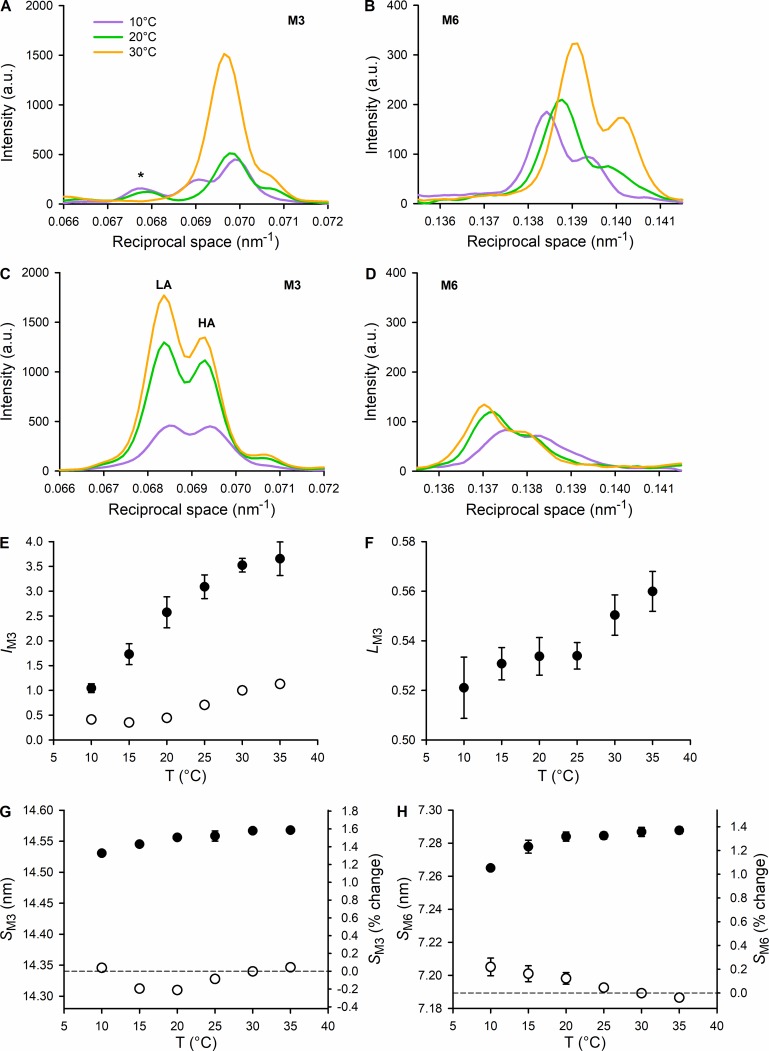
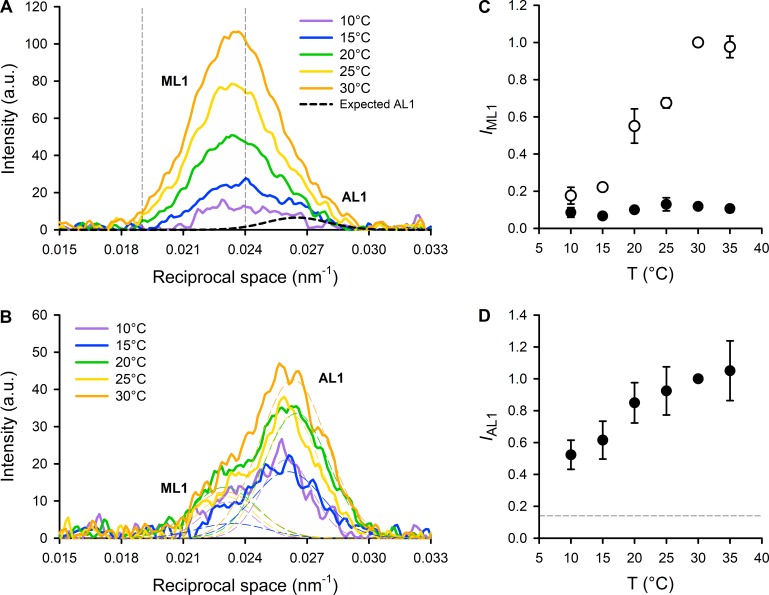
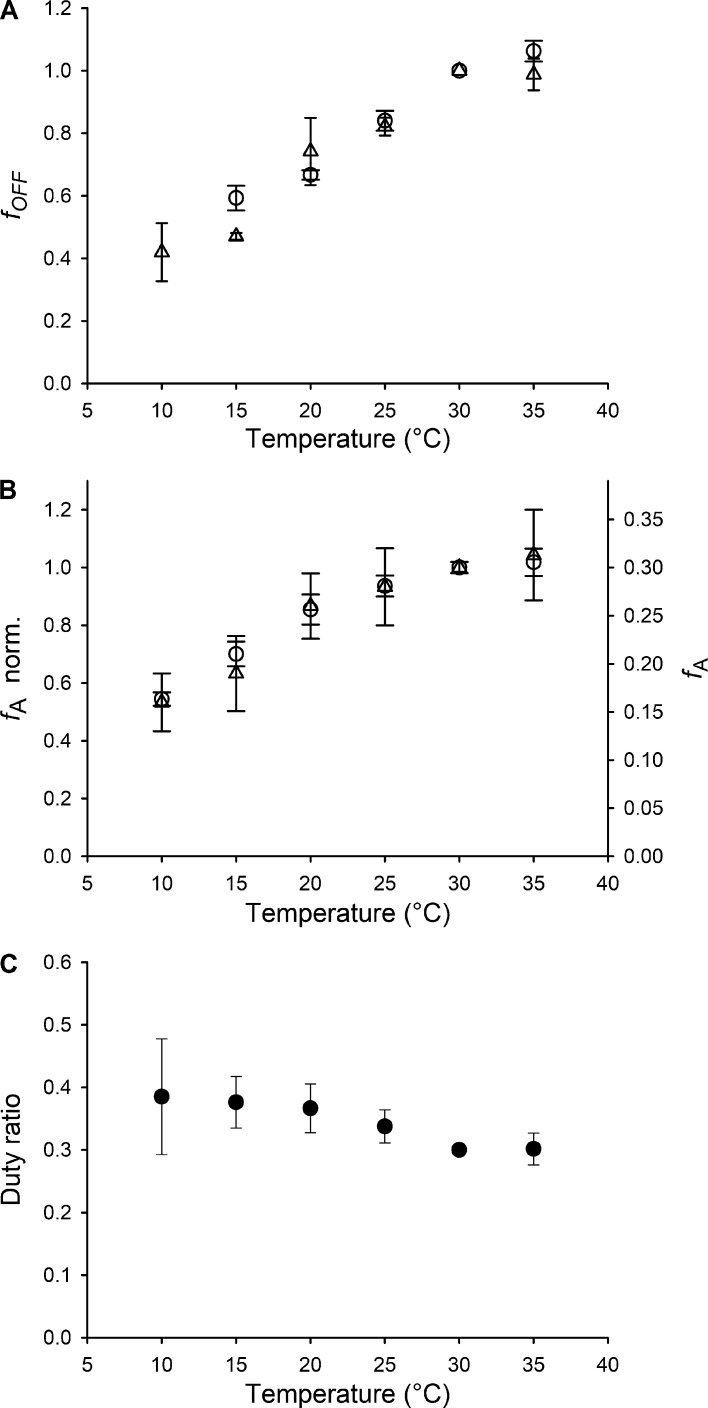
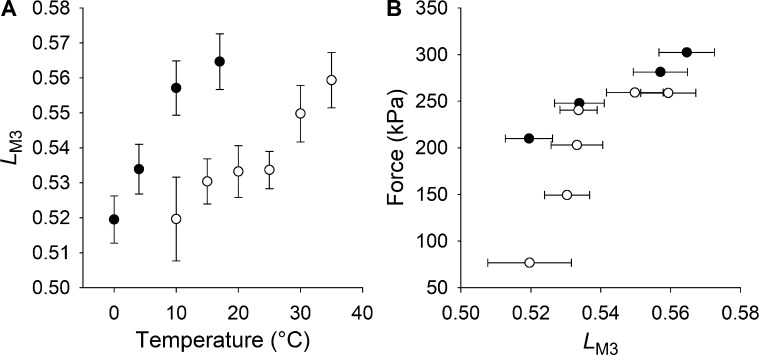
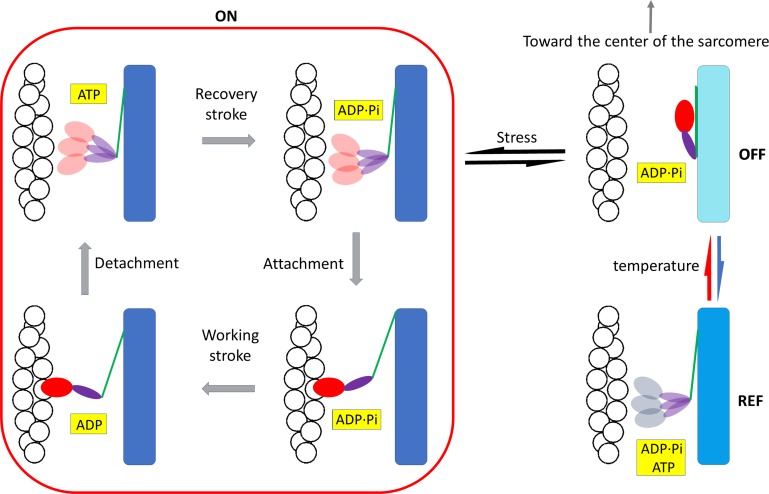
Myosin motors in the thick filament of resting striated (skeletal and cardiac) muscle are trapped in an OFF state, in which the motors are packed in helical tracks on the filament surface, inhibiting their interactions with actin and utilization of ATP. To investigate the structural changes induced in the thick filament of mammalian skeletal muscle by changes in temperature, we collected x-ray diffraction patterns from the fast skeletal muscle extensor digitorum longus of the mouse in the temperature range from near physiological (35°C) to 10°C, in which the maximal isometric force (T 0) shows a threefold decrease. In resting muscle, x-ray reflections signaling the OFF state of the thick filament indicate that cooling produces a progressive disruption of the OFF state with motors moving away from the ordered helical tracks on the surface of the thick filament. We find that the number of myosin motors in the OFF state at 10°C is half of that at 35°C. At T 0, changes in the x-ray signals that report the fraction and conformation of actin-attached motors can be explained if the threefold decrease in force associated with lowering temperature is due not only to a decrease in the force-generating transition in the actin-attached motors but also to a twofold decrease in the number of such motors. Thus, lowering the temperature reduces to the same extent the fraction of motors in the OFF state at rest and the fraction of motors attached to actin at T 0, suggesting that motors that leave the OFF state accumulate in a disordered refractory state that makes them unavailable for interaction with actin upon stimulation. This regulatory effect of temperature on the thick filament of mammalian skeletal muscle could represent an energetically convenient mechanism for hibernating animals.
© 2019 Caremani et al.
Figures
Comment in
-
Cold temperatures put a freeze on myosin activation.J Gen Physiol. 2019 Nov 4;151(11):1247. doi: 10.1085/jgp.201912509. Epub 2019 Oct 16. J Gen Physiol. 2019. PMID: 31619457 Free PMC article.
-
A case for reduced energy utilization associated with spatial disorder of myosin in skeletal muscle.J Gen Physiol. 2020 Jan 6;152(1):e201912444. doi: 10.1085/jgp.201912444. J Gen Physiol. 2020. PMID: 31740447 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
