

Circadian clock genes and the transcriptional architecture of the clock mechanism
- PMID: 31557726
- PMCID: PMC6872945
- DOI: 10.1530/JME-19-0153
Circadian clock genes and the transcriptional architecture of the clock mechanism
Abstract
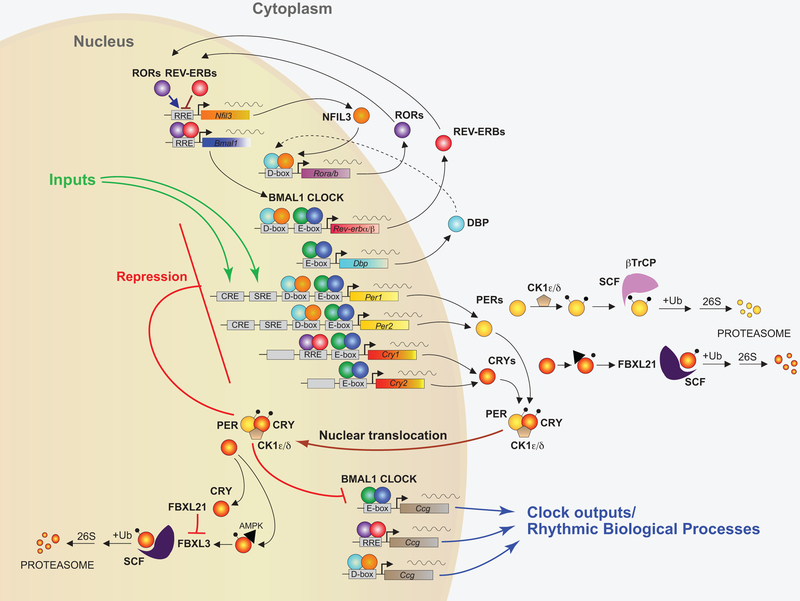
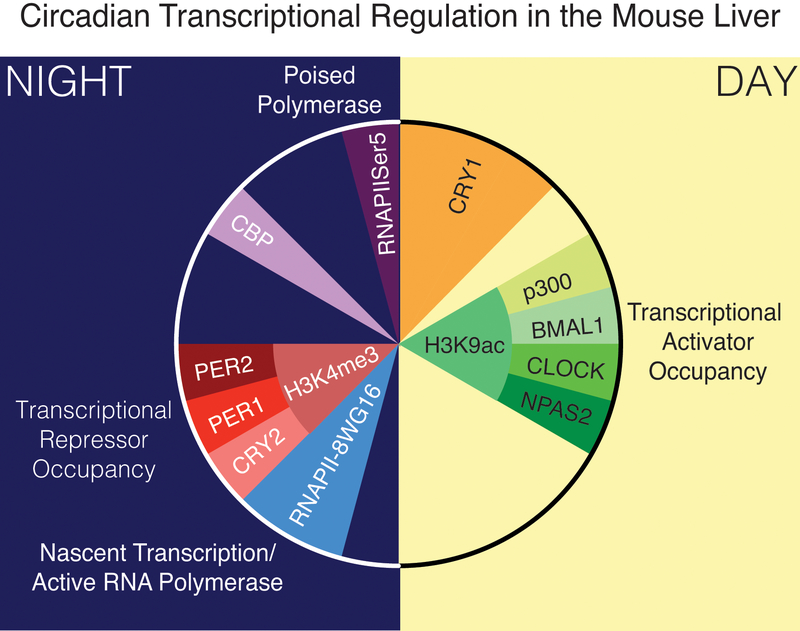
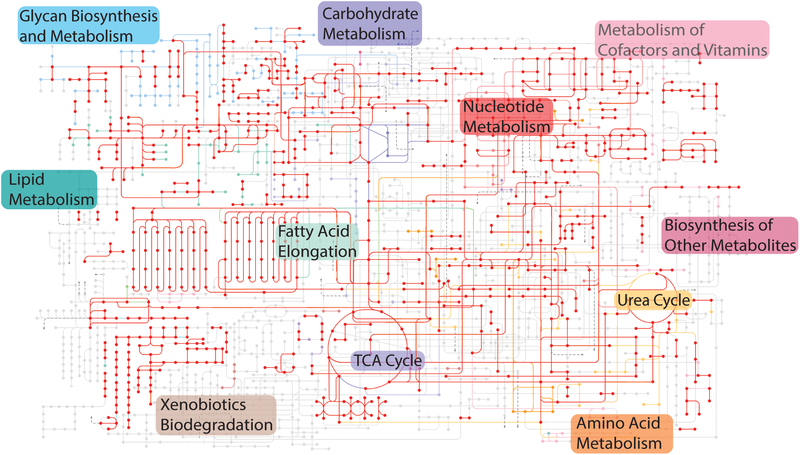
The mammalian circadian clock has evolved as an adaptation to the 24-h light/darkness cycle on earth. Maintaining cellular activities in synchrony with the activities of the organism (such as eating and sleeping) helps different tissue and organ systems coordinate and optimize their performance. The full extent of the mechanisms by which cells maintain the clock are still under investigation, but involve a core set of clock genes that regulate large networks of gene transcription both by direct transcriptional activation/repression as well as the recruitment of proteins that modify chromatin states more broadly.
Keywords: circadian clock; clock genes; mammalian; transcription.
Figures
References
-
- ABRAHAMSON EE & MOORE RY 2001. Suprachiasmatic nucleus in the mouse: retinal innervation, intrinsic organization and efferent projections. Brain Res, 916, 172–91. - PubMed
-
- ALBRECHT U 2012. Timing to perfection: the biology of central and peripheral circadian clocks. Neuron, 74, 246–60. - PubMed
-
- ASHER G, GATFIELD D, STRATMANN M, REINKE H, DIBNER C, KREPPEL F, MOSTOSLAVSKY R, ALT FW & SCHIBLER U 2008. SIRT1 regulates circadian clock gene expression through PER2 deacetylation. Cell, 134, 317–28. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
