

Dynamic changes in epithelial cell morphology control thymic organ size during atrophy and regeneration
- PMID: 31562306
- PMCID: PMC6765001
- DOI: 10.1038/s41467-019-11879-2
Dynamic changes in epithelial cell morphology control thymic organ size during atrophy and regeneration
Abstract
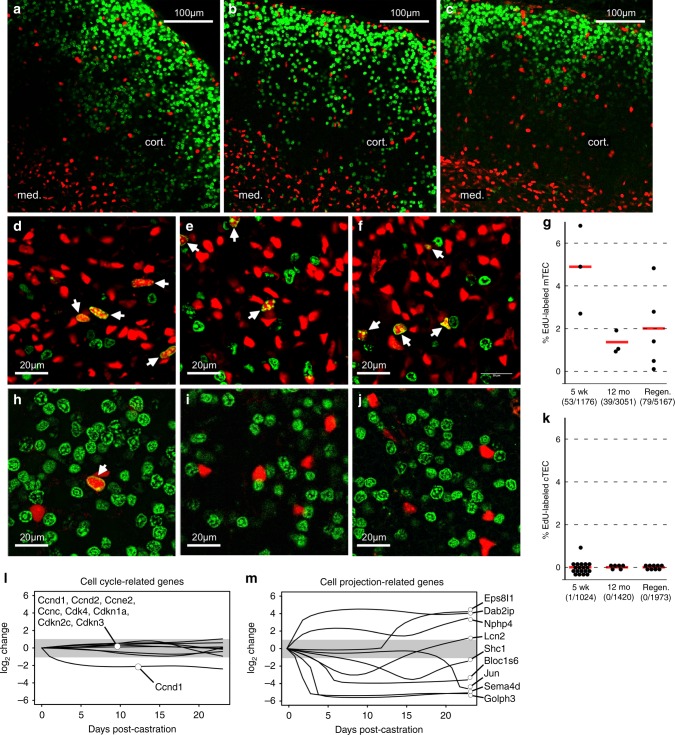
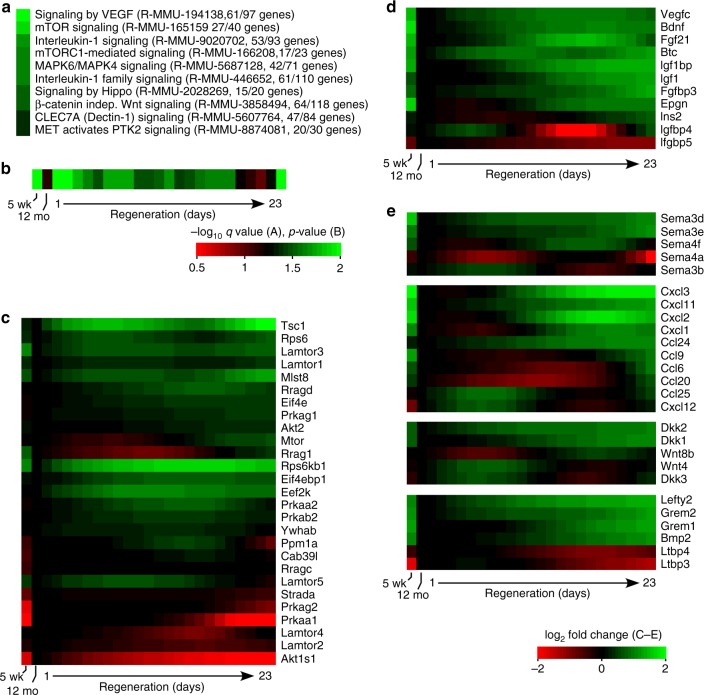
T lymphocytes must be produced throughout life, yet the thymus, where T lymphocytes are made, exhibits accelerated atrophy with age. Even in advanced atrophy, however, the thymus remains plastic, and can be regenerated by appropriate stimuli. Logically, thymic atrophy is thought to reflect senescent cell death, while regeneration requires proliferation of stem or progenitor cells, although evidence is scarce. Here we use conditional reporters to show that accelerated thymic atrophy reflects contraction of complex cell projections unique to cortical epithelial cells, while regeneration requires their regrowth. Both atrophy and regeneration are independent of changes in epithelial cell number, suggesting that the size of the thymus is regulated primarily by rate-limiting morphological changes in cortical stroma, rather than by their cell death or proliferation. Our data also suggest that cortical epithelial morphology is under the control of medullary stromal signals, revealing a previously unrecognized endocrine-paracrine signaling axis in the thymus.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Up-regulation of IL-7, stromal-derived factor-1 alpha, thymus-expressed chemokine, and secondary lymphoid tissue chemokine gene expression in the stromal cells in response to thymocyte depletion: implication for thymus reconstitution.J Immunol. 2005 Aug 15;175(4):2321-30. doi: 10.4049/jimmunol.175.4.2321. J Immunol. 2005. PMID: 16081802
-
Persistent degenerative changes in thymic organ function revealed by an inducible model of organ regrowth.Aging Cell. 2012 Feb;11(1):169-77. doi: 10.1111/j.1474-9726.2011.00773.x. Epub 2011 Dec 28. Aging Cell. 2012. PMID: 22103718 Free PMC article.
-
FSP1(+) fibroblast subpopulation is essential for the maintenance and regeneration of medullary thymic epithelial cells.Sci Rep. 2015 Oct 8;5:14871. doi: 10.1038/srep14871. Sci Rep. 2015. PMID: 26445893 Free PMC article.
-
Thymic epithelial progenitor cells and thymus regeneration: an update.Cell Res. 2007 Jan;17(1):50-5. doi: 10.1038/sj.cr.7310114. Cell Res. 2007. PMID: 17146450 Review.
-
Thymic stromal cells: Roles in atrophy and age-associated dysfunction of the thymus.Exp Gerontol. 2018 May;105:113-117. doi: 10.1016/j.exger.2017.12.022. Epub 2017 Dec 24. Exp Gerontol. 2018. PMID: 29278750 Free PMC article. Review.
Cited by
-
Androgen Receptors in Epithelial Cells Regulate Thymopoiesis and Recent Thymic Emigrants in Male Mice.Front Immunol. 2020 Jun 29;11:1342. doi: 10.3389/fimmu.2020.01342. eCollection 2020. Front Immunol. 2020. PMID: 32714327 Free PMC article.
-
Reduction of endocytosis and EGFR signaling is associated with the switch from isolated to clustered apoptosis during epithelial tissue remodeling in Drosophila.PLoS Biol. 2024 Oct 14;22(10):e3002823. doi: 10.1371/journal.pbio.3002823. eCollection 2024 Oct. PLoS Biol. 2024. PMID: 39401187 Free PMC article.
-
Abrogation of Notch Signaling in Embryonic TECs Impacts Postnatal mTEC Homeostasis and Thymic Involution.Front Immunol. 2022 May 30;13:867302. doi: 10.3389/fimmu.2022.867302. eCollection 2022. Front Immunol. 2022. PMID: 35707539 Free PMC article.
-
Age-related thymic involution: Mechanisms and functional impact.Aging Cell. 2022 Aug;21(8):e13671. doi: 10.1111/acel.13671. Epub 2022 Jul 12. Aging Cell. 2022. PMID: 35822239 Free PMC article. Review.
-
Immune tolerance and the prevention of autoimmune diseases essentially depend on thymic tissue homeostasis.Front Immunol. 2024 Mar 20;15:1339714. doi: 10.3389/fimmu.2024.1339714. eCollection 2024. Front Immunol. 2024. PMID: 38571951 Free PMC article. Review.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
