

RBX1 prompts degradation of EXO1 to limit the homologous recombination pathway of DNA double-strand break repair in G1 phase
- PMID: 31562368
- PMCID: PMC7205894
- DOI: 10.1038/s41418-019-0424-4
RBX1 prompts degradation of EXO1 to limit the homologous recombination pathway of DNA double-strand break repair in G1 phase
Abstract
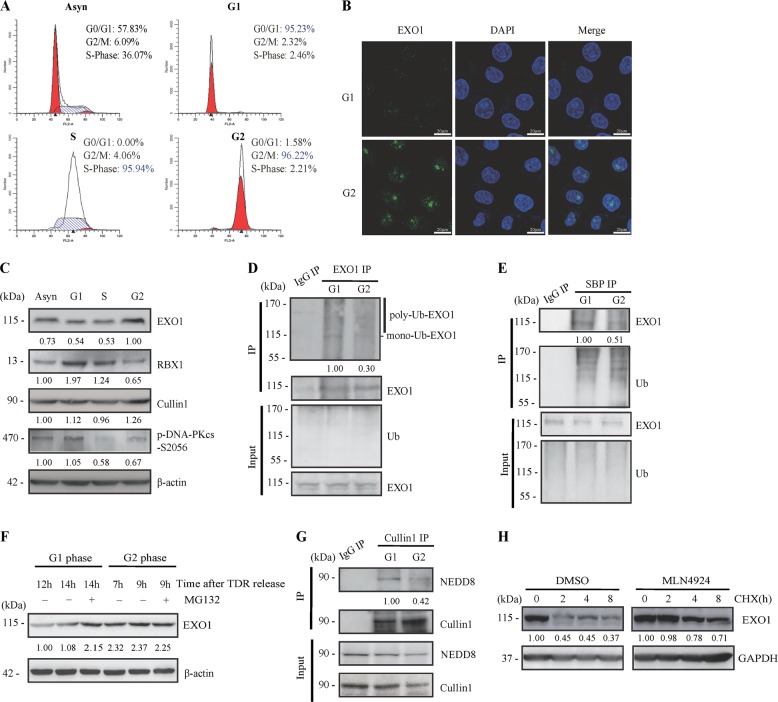
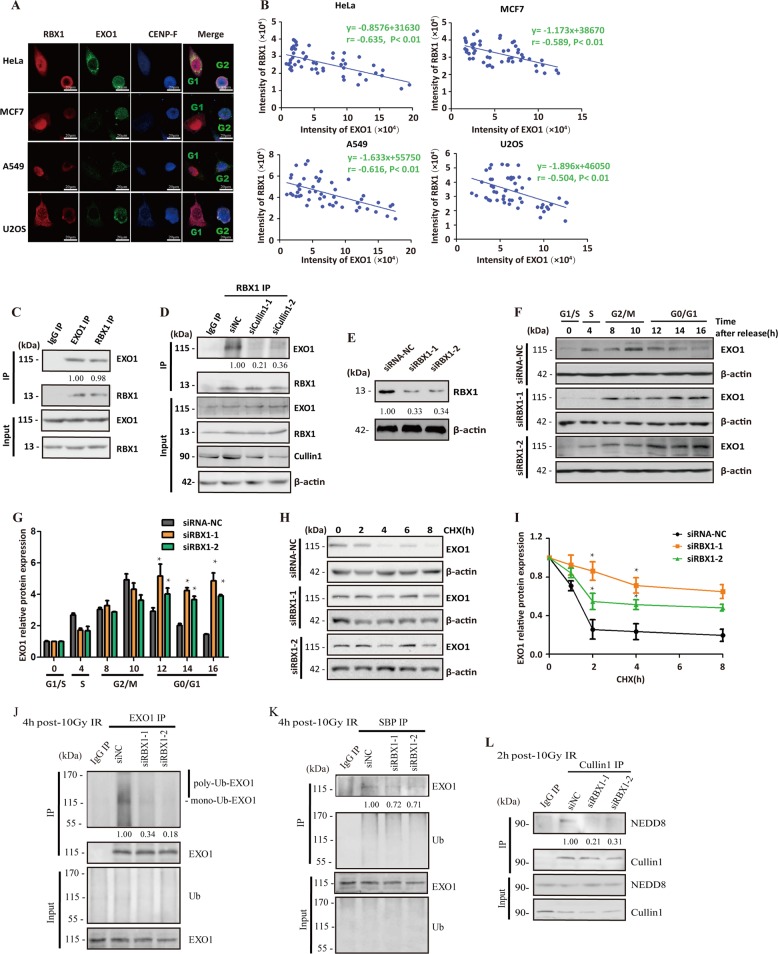
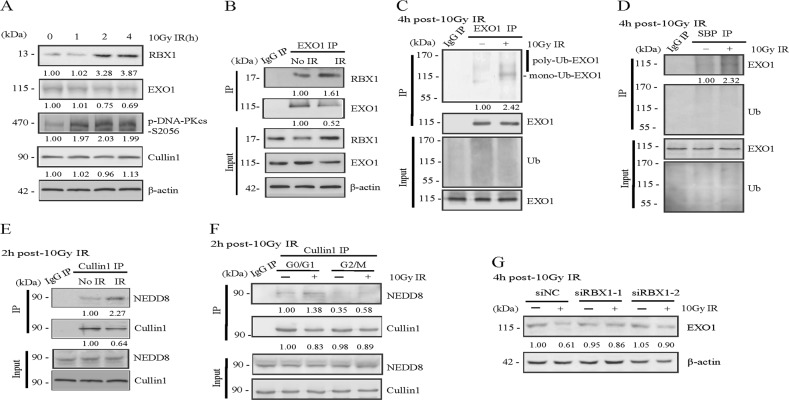
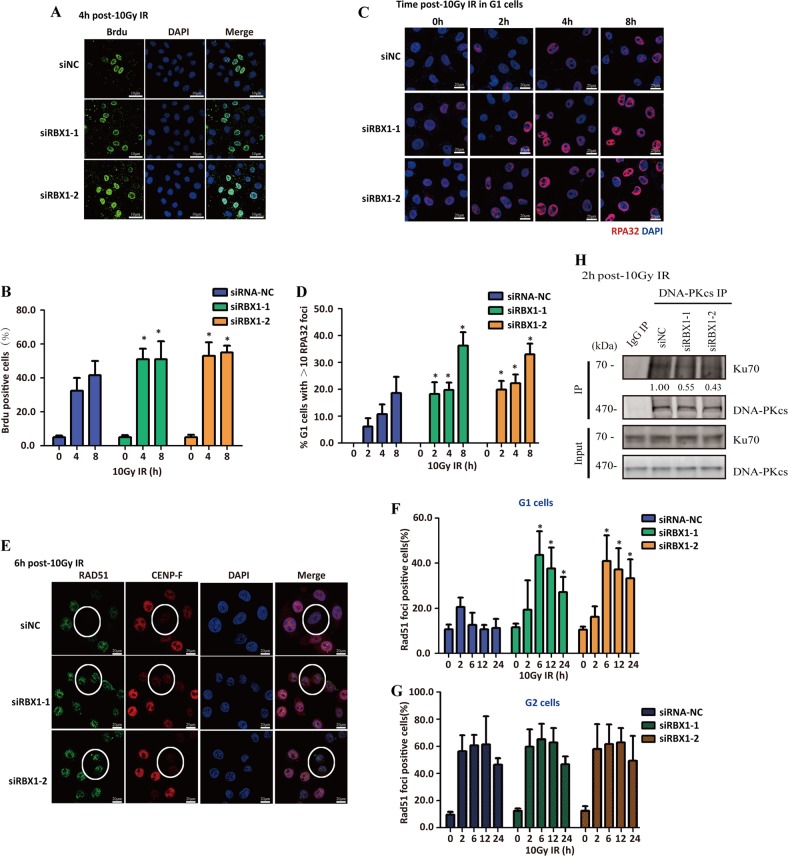
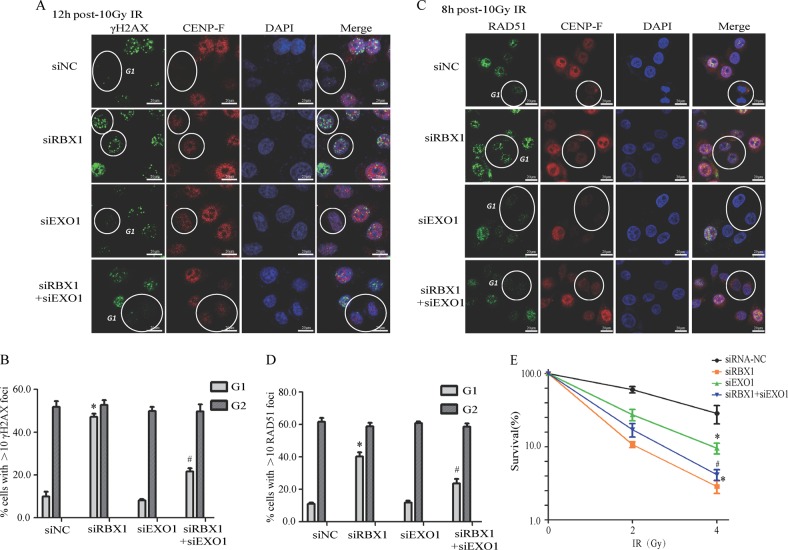
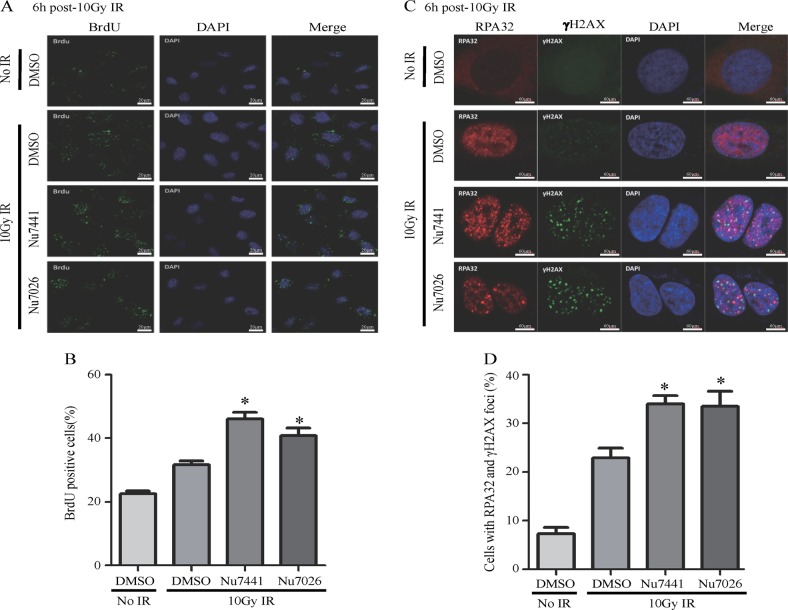
End resection of DNA double-strand breaks (DSBs) to form 3' single-strand DNA (ssDNA) is critical to initiate the homologous recombination (HR) pathway of DSB repair. HR pathway is strictly limited in the G1-phase cells because of lack of homologous DNA as the templates. Exonuclease 1 (EXO1) is the key molecule responsible for 3' ssDNA formation of DSB end resection. We revealed that EXO1 is inactivated in G1-phase cells via ubiquitination-mediated degradation, resulting from an elevated expression level of RING-box protein 1 (RBX1) in G1 phase. The increased RBX1 significantly prompted the neddylation of Cullin1 and contributed to the G1 phase-specific degradation of EXO1. Knockdown of RBX1 remarkedly attenuated the degradation of EXO1 and increased the end resection and HR activity in γ-irradiated G1-phase cells, as demonstrated by the increased formation of RPA32, BrdU, and RAD51 foci. And EXO1 depletion mitigated DNA repair defects due to RBX1 reduction. Moreover, increased autophosphorylation of DNA-PKcs at S2056 was found to be responsible for the higher expression level of the RBX1 in the G1 phase. Inactivation of DNA-PKcs decreased RBX1 expression, and simultaneously increased EXO1 expression and DSB end resection in G1-phase cells. This study demonstrates a new mechanism for restraining the HR pathway of DNA DSB repair in G1 phase via RBX1-prompted inactivation of EXO1.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
