

The roles of fission yeast exonuclease 5 in nuclear and mitochondrial genome stability
- PMID: 31563844
- PMCID: PMC6990415
- DOI: 10.1016/j.dnarep.2019.102720
The roles of fission yeast exonuclease 5 in nuclear and mitochondrial genome stability
Abstract
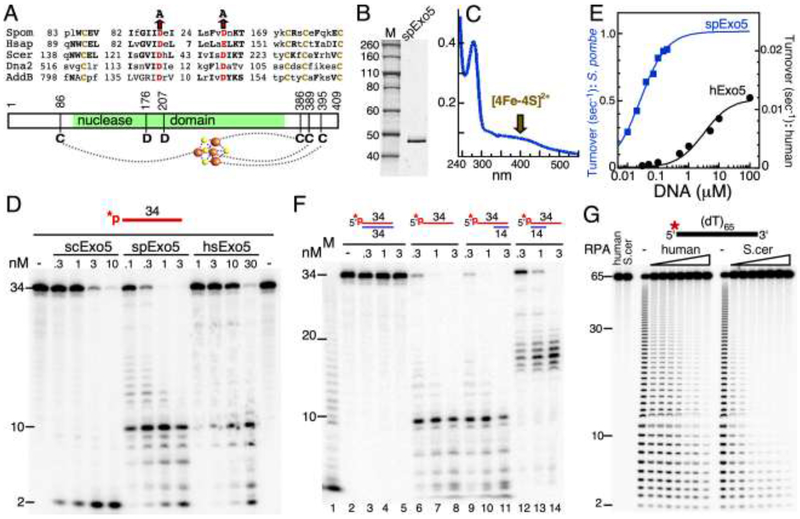
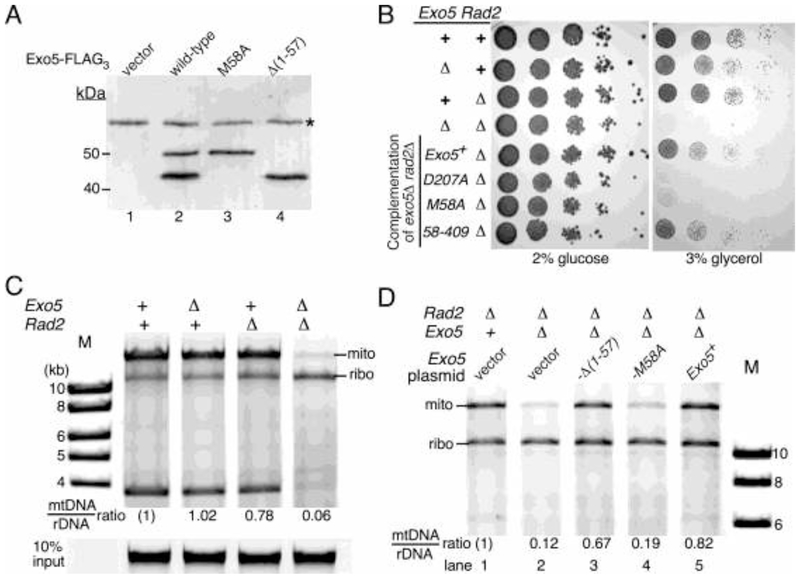
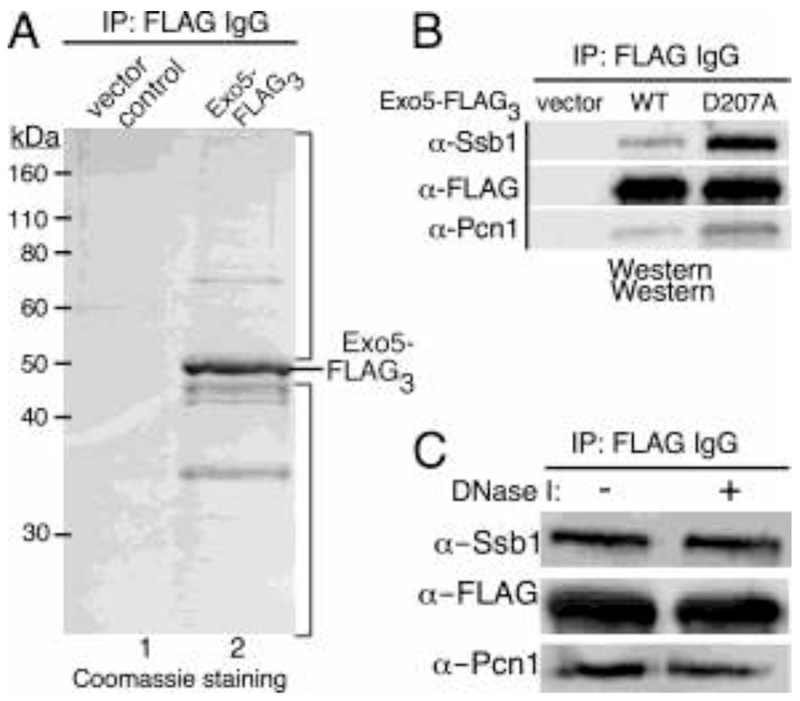
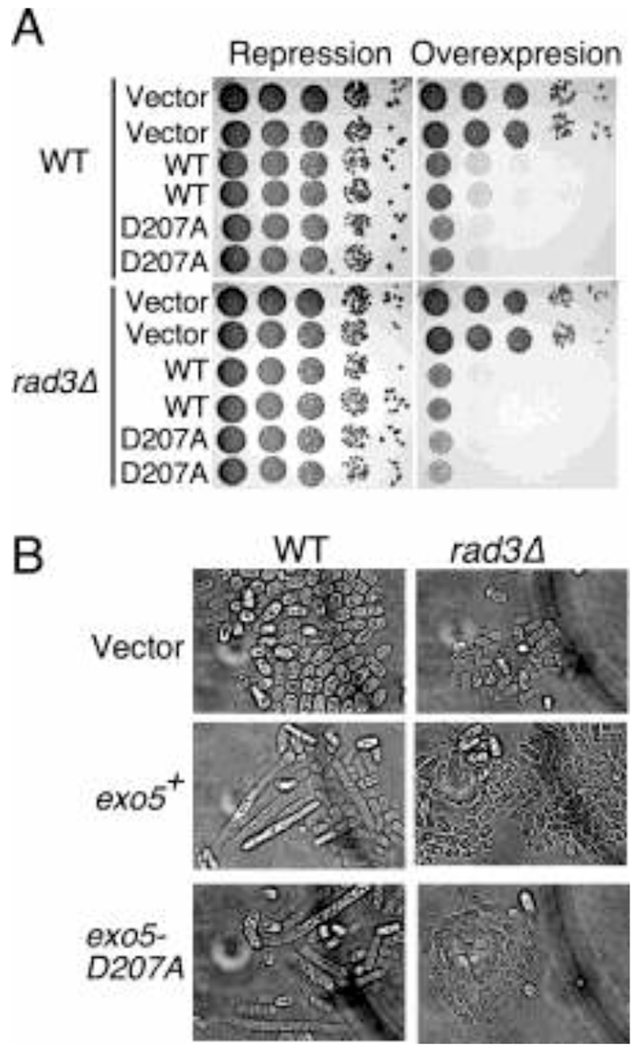
The Exo5 family consists of bi-directional, single-stranded DNA-specific exonucleases that contain an iron-sulfur cluster as a structural motif and have multiple roles in DNA metabolism. S. cerevisiae Exo5 is essential for mitochondrial genome maintenance, while the human ortholog is important for nuclear genome stability and DNA repair. Here, we identify the Exo5 ortholog in Schizosaccharomyes pombe (spExo5). The activity of spExo5 is highly similar to that of the human enzyme. When the single-stranded DNA is coated with single-stranded DNA binding protein RPA, spExo5 become a 5'-specific exonuclease. Exo5Δ mutants are sensitive to various DNA damaging agents, particularly interstrand crosslinking agents. An epistasis analysis places exo5+ in the Fanconi pathway for interstrand crosslink repair. Exo5+ is in a redundant pathway with rad2+, which encodes the flap endonuclease FEN1, for mitochondrial genome maintenance. Deletion of both genes lead to severe depletion of the mitochondrial genome, and defects in respiration, indicating that either spExo5 or spFEN1 is necessary for mitochondrial DNA metabolism.
Keywords: Exo5; Exonuclease; Fission yeast; Interstrand crosslink repair; Mitochondrial replication.
Copyright © 2019 Elsevier B.V. All rights reserved.
Figures

References
-
- Burgers PMJ, Bauer GA, Tam L, Exonuclease V from Saccharomyces cerevisiae. A 5′-- -3′-deoxyribonuclease that produces dinucleotides in a sequential fashion, J. Biol. Chem, 263 (1988) 8099–8105. - PubMed
-
- Paumard-Hernandez B, Calvete O, Inglada Perez L, Tejero H, Al-Shahrour F, Pita G, Barroso A, Carlos Trivino J, Urioste M, Valverde C, Gonzalez Billalabeitia E, Quiroga V, Francisco Rodriguez Moreno J, Fernandez Aramburo A, Lopez C, Maroto P, Sastre J, Jose Juan Fita M, Duran I, Lorenzo-Lorenzo I, Iranzo P, Garcia Del Muro X, Ros S, Zambrana F, Maria Autran A, Benitez J, Whole exome sequencing identifies PLEC, EXO5 and DNAH7 as novel susceptibility genes in testicular cancer, Int J Cancer, 143 (2018) 1954–1962. - PubMed
-
- Knijnenburg TA, Wang L, Zimmermann MT, Chambwe N, Gao GF, Cherniack AD, Fan H, Shen H, Way GP, Greene CS, Liu Y, Akbani R, Feng B, Donehower LA, Miller C, Shen Y, Karimi M, Chen H, Kim P, Jia P, Shinbrot E, Zhang S, Liu J, Hu H, Bailey MH, Yau C, Wolf D, Zhao Z, Weinstein JN, Li L, Ding L, Mills GB, Laird PW, Wheeler DA, Shmulevich I, Cancer N Genome Atlas Research, Monnat RJ Jr., Xiao Y, Wang C, Genomic and Molecular Landscape of DNA Damage Repair Deficiency across The Cancer Genome Atlas, Cell Rep, 23 (2018) 239–254 e236. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
