

Shared Song Detector Neurons in Drosophila Male and Female Brains Drive Sex-Specific Behaviors
- PMID: 31564492
- PMCID: PMC6885007
- DOI: 10.1016/j.cub.2019.08.008
Shared Song Detector Neurons in Drosophila Male and Female Brains Drive Sex-Specific Behaviors
Erratum in
-
Shared Song Detector Neurons in Drosophila Male and Female Brains Drive Sex-Specific Behaviors.Curr Biol. 2023 Sep 11;33(17):3796-3800. doi: 10.1016/j.cub.2023.08.006. Curr Biol. 2023. PMID: 37699336 No abstract available.
Abstract
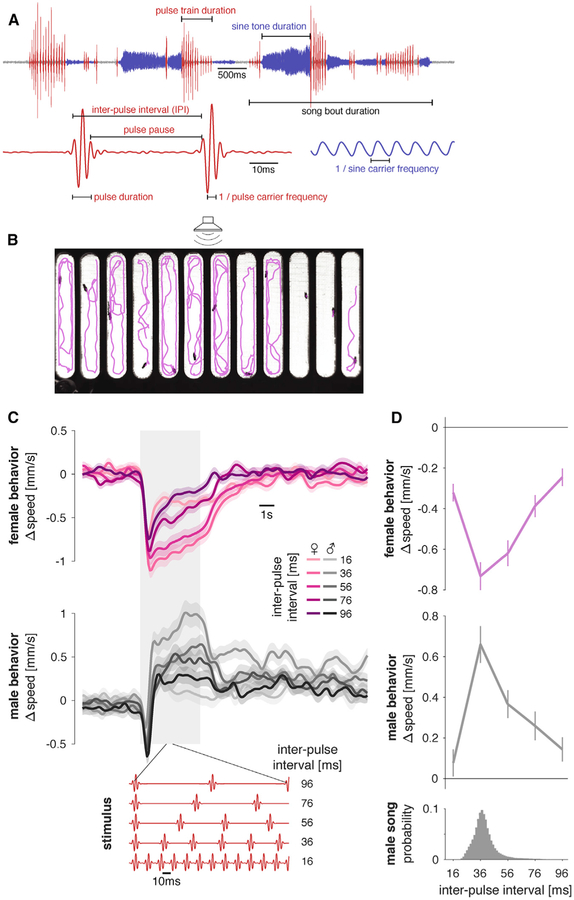
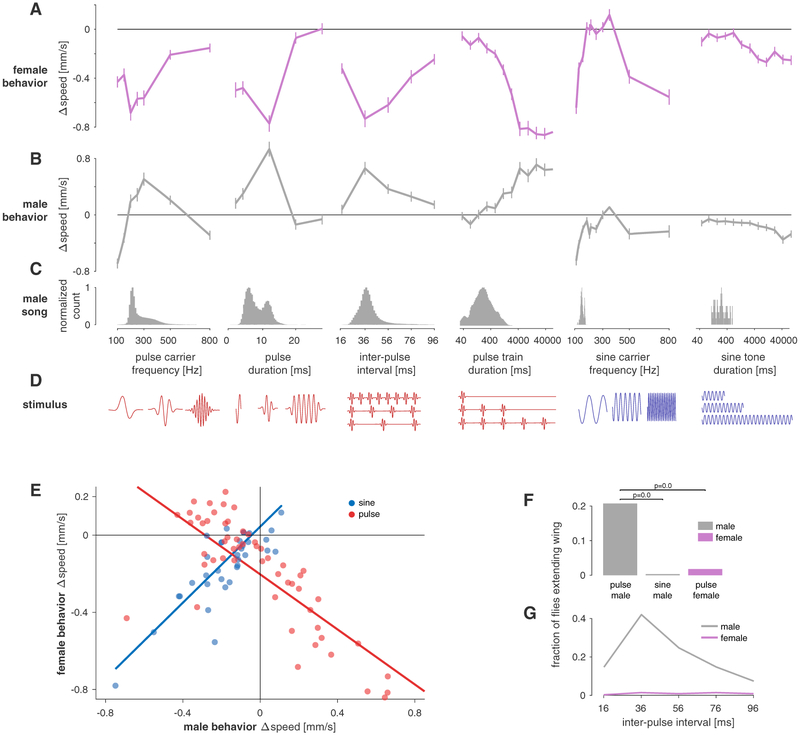
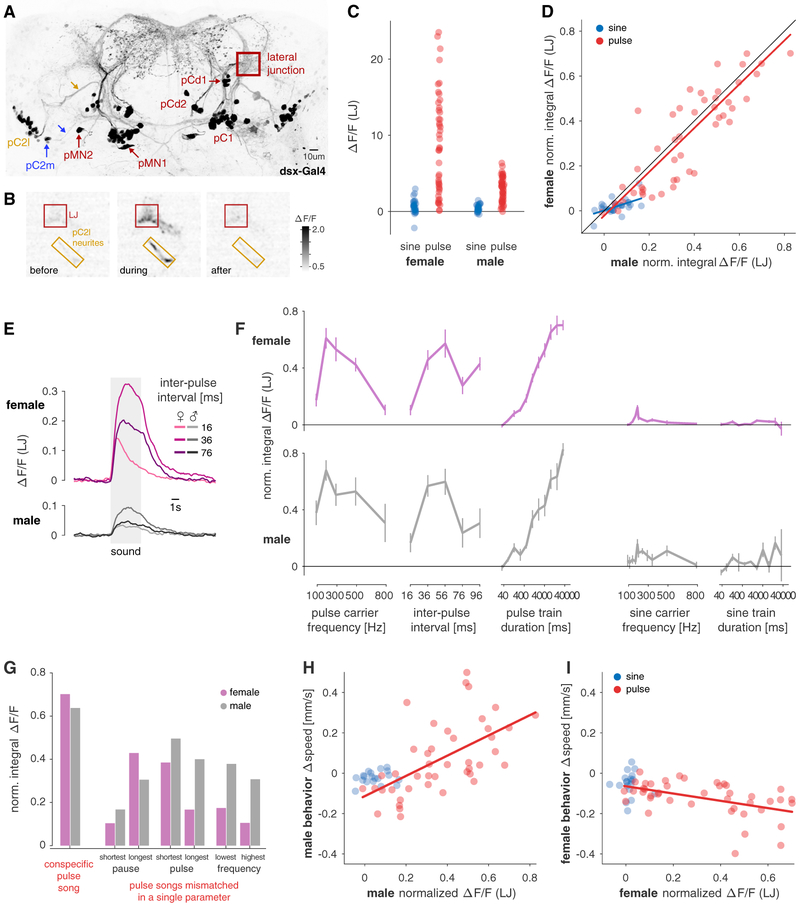
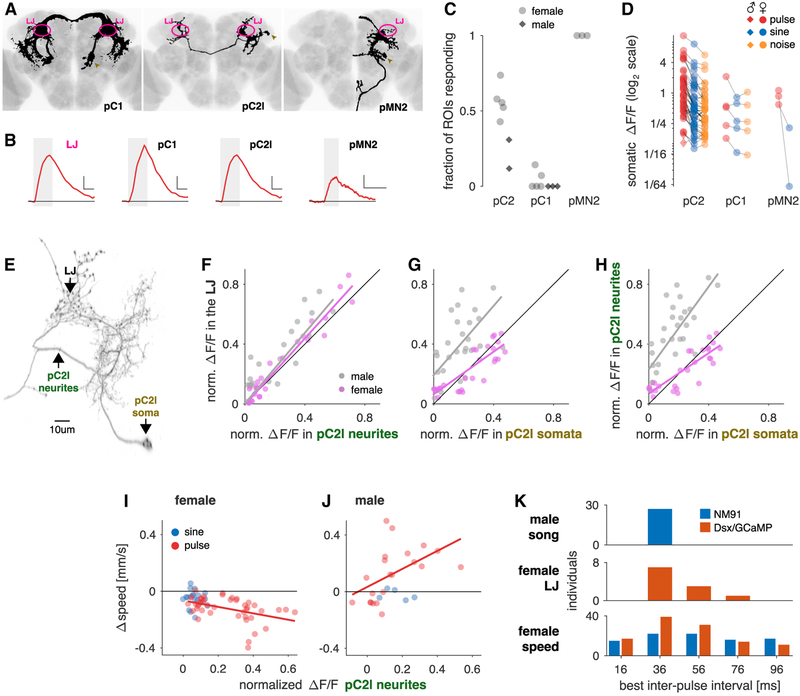
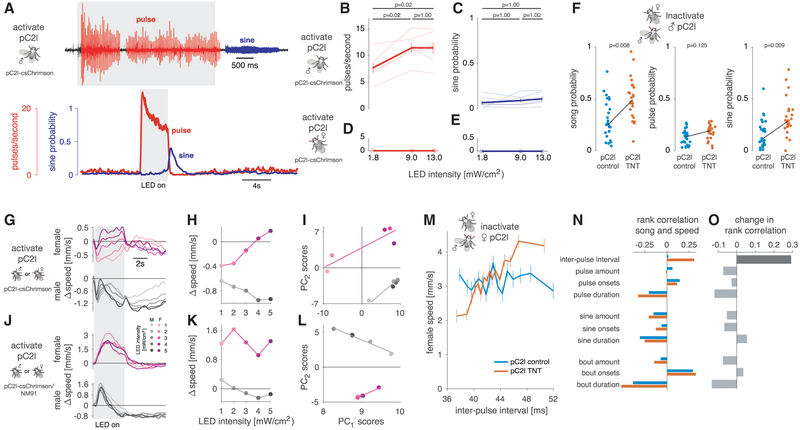
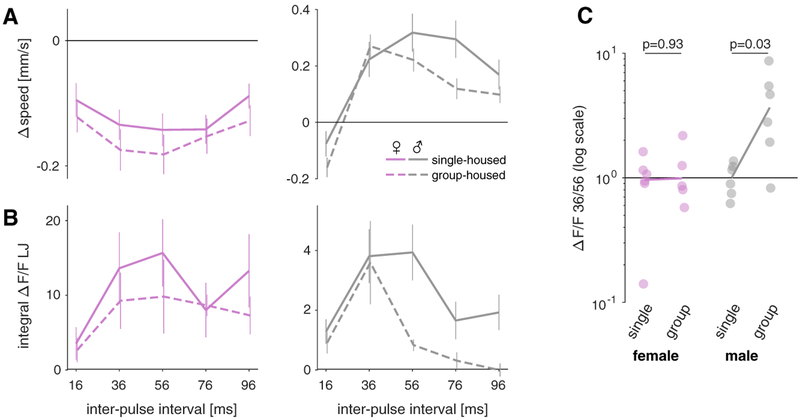
Males and females often produce distinct responses to the same sensory stimuli. How such differences arise-at the level of sensory processing or in the circuits that generate behavior-remains largely unresolved across sensory modalities. We address this issue in the acoustic communication system of Drosophila. During courtship, males generate time-varying songs, and each sex responds with specific behaviors. We characterize male and female behavioral tuning for all aspects of song and show that feature tuning is similar between sexes, suggesting sex-shared song detectors drive divergent behaviors. We then identify higher-order neurons in the Drosophila brain, called pC2, that are tuned for multiple temporal aspects of one mode of the male's song and drive sex-specific behaviors. We thus uncover neurons that are specifically tuned to an acoustic communication signal and that reside at the sensory-motor interface, flexibly linking auditory perception with sex-specific behavioral responses.
Keywords: Drosophila; acoustic communication; auditory; behavior; courtship; neural circuits; pattern recognition; sensorimotor transformation; sexually dimorphic; social experience; song detection.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
References
-
- Dulac C, and Wagner S (2006). Genetic Analysis of Brain Circuits Underlying Pheromone Signaling. Annu. Rev. Genet 40, 449–467. - PubMed
-
- Kurtovic A, Widmer A, and Dickson BJ (2007). A single class of olfactory neurons mediates behavioural responses to a Drosophila sex pheromone. Nature 446, 542–546. - PubMed
-
- Billeter J-C, Atallah J, Krupp JJ, Millar JG, and Levine JD (2009). Specialized cells tag sexual and species identity in Drosophila melanogaster. Nature 461, 987–991. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
